

Mutations and rearrangements in the genome of Sulfolobus solfataricus P2
- PMID: 16740926
- PMCID: PMC1482960
- DOI: 10.1128/JB.00061-06
Mutations and rearrangements in the genome of Sulfolobus solfataricus P2
Abstract
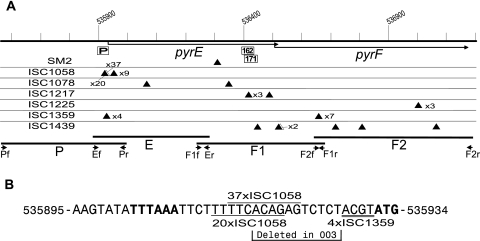
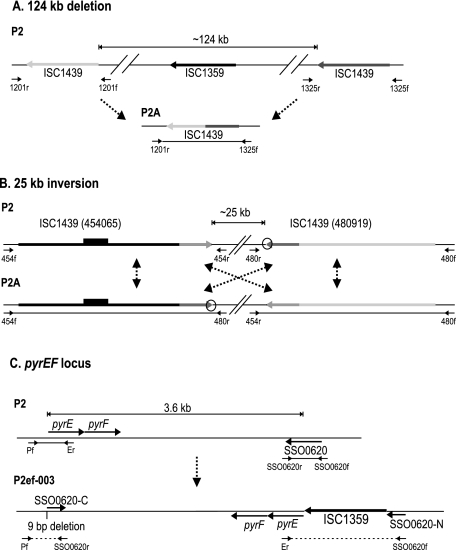
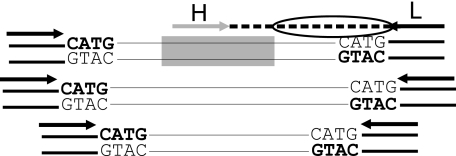
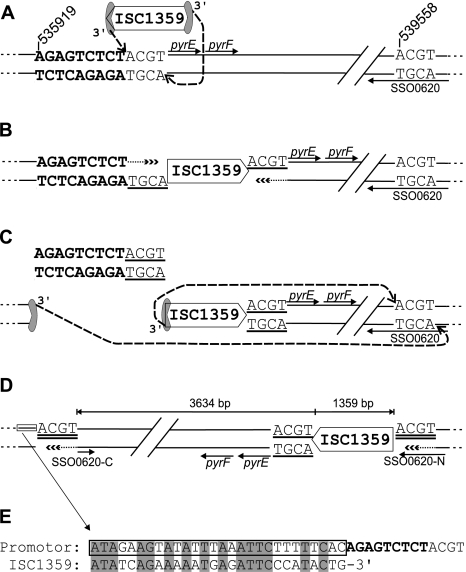
The genome of Sulfolobus solfataricus P2 carries a larger number of transposable elements than any other sequenced genome from an archaeon or bacterium and, as a consequence, may be particularly susceptible to rearrangement and change. In order to gain more insight into the natures and frequencies of different types of mutation and possible rearrangements that can occur in the genome, the pyrEF locus was examined for mutations that were isolated after selection with 5-fluoroorotic acid. About two-thirds of the 130 mutations resulted from insertions of mobile elements, including insertion sequence (IS) elements and a single nonautonomous mobile element, SM2. For each of these, the element was identified and shown to be present at its original genomic position, consistent with a progressive increase in the copy numbers of the mobile elements. In addition, several base pair substitutions, as well as small deletions, insertions, and a duplication, were observed, and about one-fifth of the mutations occurred elsewhere in the genome, possibly in an orotate transporter gene. One mutant exhibited a 5-kb genomic rearrangement at the pyrEF locus involving a two-step IS element-dependent reaction, and its boundaries were defined using a specially developed "in vitro library" strategy. Moreover, while searching for the donor mobile elements, evidence was found for two major changes that had occurred in the genome of strain P2, one constituting a single deletion of about 4% of the total genome (124 kb), while the other involved the inversion of a 25-kb region. Both were bordered by IS elements and were inferred to have arisen through recombination events. The results underline the caution required in working experimentally with an organism such as S. solfataricus with a continually changing genome.
Figures

References
-
- Blount, Z. D., and D. W. Grogan. 2005. New insertion sequences of Sulfolobus: functional properties and implications for genome evolution in hyperthermophilic archaea. Mol. Microbiol. 55:312-325. - PubMed
-
- Brügger, K., P. Redder, Q. She, F. Confalonieri, Y. Zivanovic, and R. A. Garrett. 2002. Mobile elements in archaeal genomes. FEMS Microbiol. Lett. 206:131-141. - PubMed
-
- Brügger, K., E. Torarinsson, P. Redder, L. Chen, and R. A. Garrett. 2004. Shuffling of Sulfolobus genomes by autonomous and non-autonomous mobile elements. Biochem. Soc. Trans. 32:179-183. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
