

Borrelia burgdorferi EbfC, a novel, chromosomally encoded protein, binds specific DNA sequences adjacent to erp loci on the spirochete's resident cp32 prophages
- PMID: 16740939
- PMCID: PMC1482946
- DOI: 10.1128/JB.00005-06
Borrelia burgdorferi EbfC, a novel, chromosomally encoded protein, binds specific DNA sequences adjacent to erp loci on the spirochete's resident cp32 prophages
Abstract
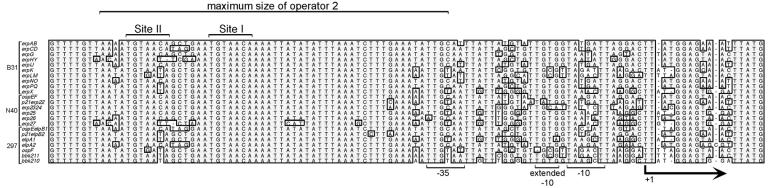
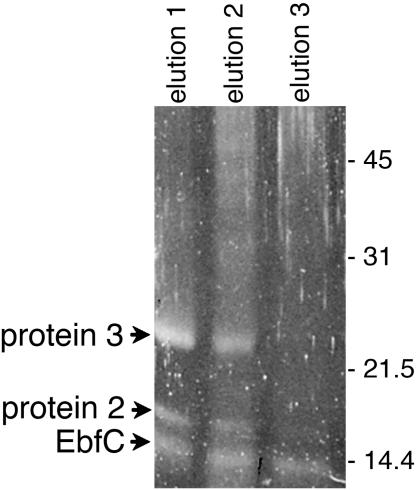
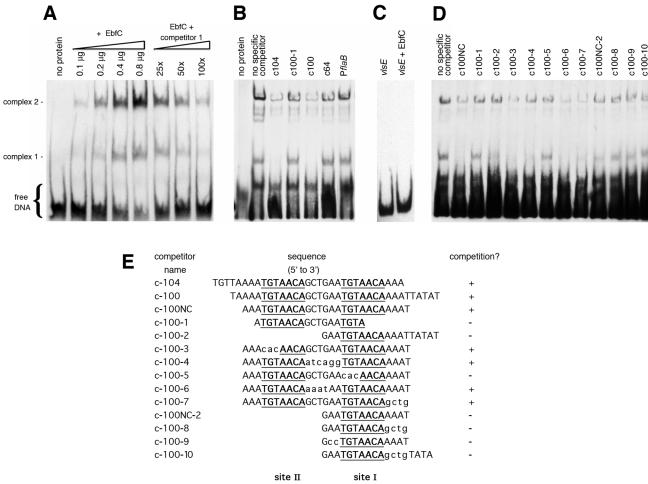
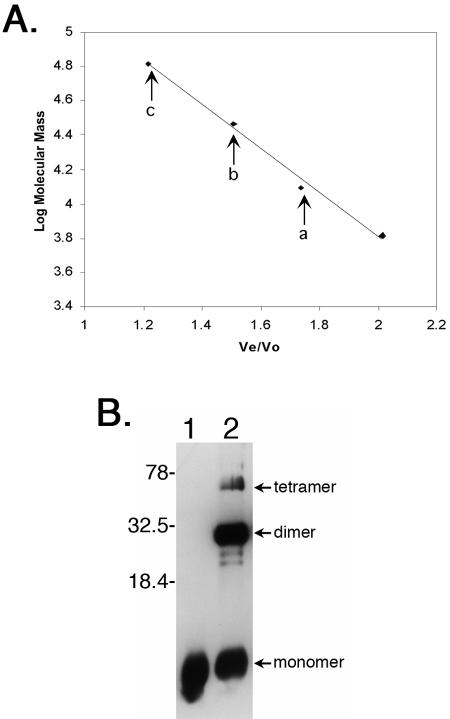
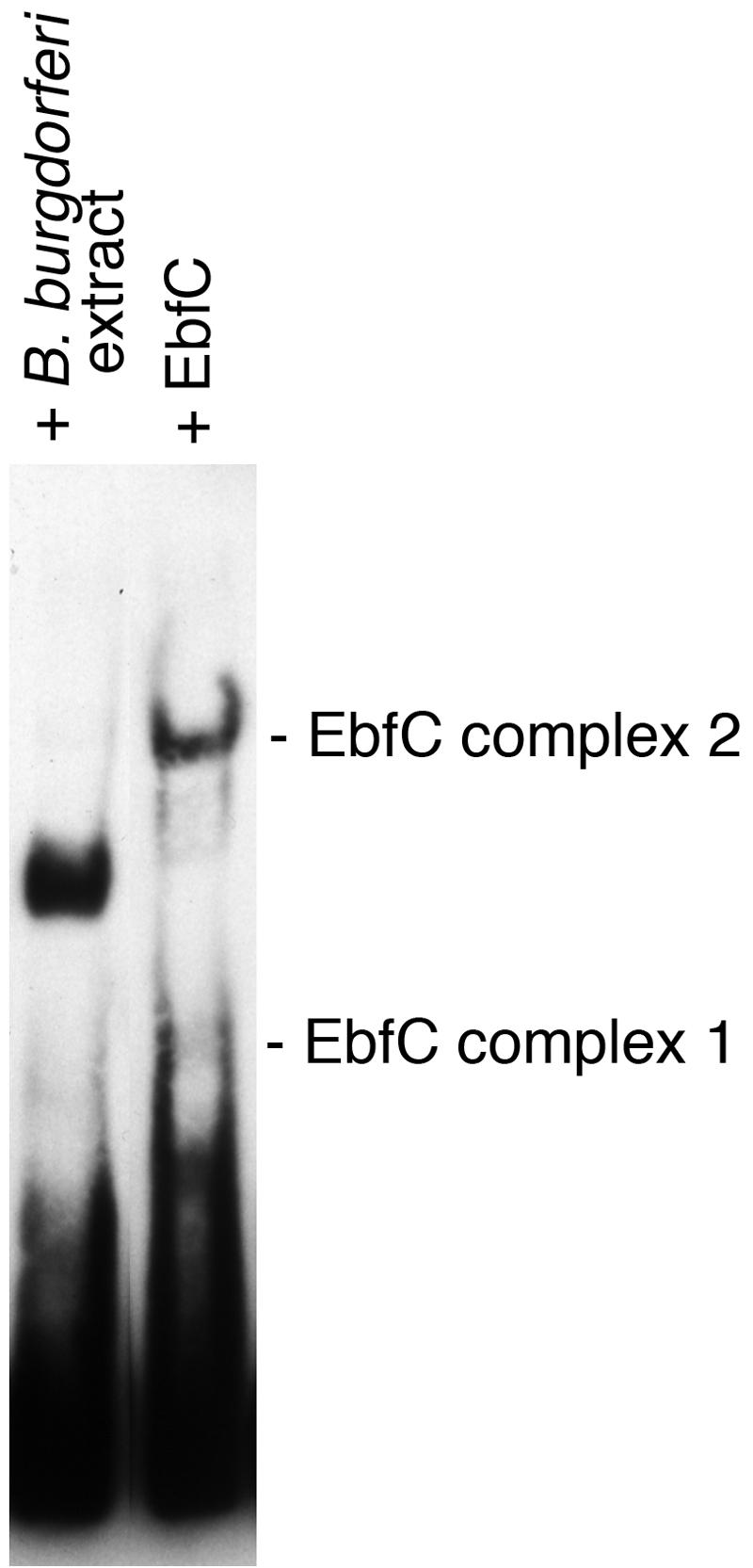
All examined isolates of the Lyme disease spirochete, Borrelia burgdorferi, naturally maintain numerous variants of a prophage family as circular cp32 episomes. Each cp32 carries a locus encoding one or two different Erp outer membrane, surface-exposed lipoproteins. Many of the Erp proteins bind a host complement regulator, factor H, which is hypothesized to protect the spirochete from complement-mediated killing. We now describe the isolation and characterization of a novel, chromosomally encoded protein, EbfC, that binds specific DNA sequences located immediately 5' of all erp loci. This is one of the first site-specific DNA-binding proteins to be identified in any spirochete. The location of the ebfC gene on the B. burgdorferi chromosome suggests that the cp32 prophages have evolved to use this bacterial host protein for their own benefit and that EbfC probably plays additional roles in the bacterium. A wide range of other bacteria encode homologs of EbfC, none of which have been well characterized, so demonstration that B. burgdorferi EbfC is a site-specific DNA-binding protein has broad implications across the eubacterial kingdom.
Figures

References
-
- Akins, D. R., S. F. Porcella, T. G. Popova, D. Shevchenko, S. I. Baker, M. Li, M. V. Norgard, and J. D. Radolf. 1995. Evidence for in vivo but not in vitro expression of a Borrelia burgdorferi outer surface protein F (OspF) homologue. Mol. Microbiol. 18:507-520. - PubMed
-
- Alitalo, A., T. Meri, H. Lankinen, I. Seppälä, P. Lahdenne, P. S. Hefty, D. Akins, and S. Meri. 2002. Complement inhibitor factor H binding to Lyme disease spirochetes is mediated by inducible expression of multiple plasmid-encoded outer surface protein E paralogs. J. Immunol. 169:3847-3853. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
