

Microfibrillar structure of type I collagen in situ
- PMID: 16751282
- PMCID: PMC1473175
- DOI: 10.1073/pnas.0502718103
Microfibrillar structure of type I collagen in situ
Abstract
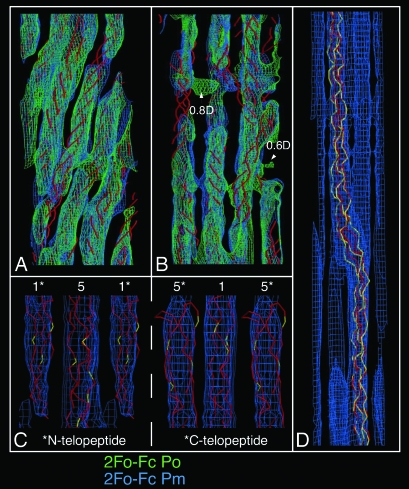
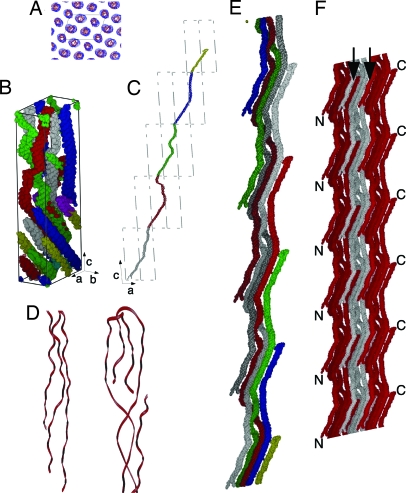
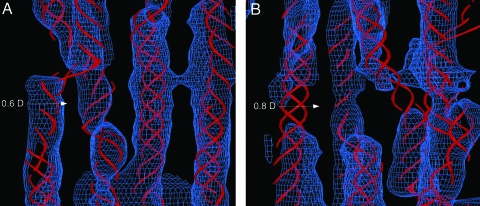
The fibrous collagens are ubiquitous in animals and form the structural basis of all mammalian connective tissues, including those of the heart, vasculature, skin, cornea, bones, and tendons. However, in comparison with what is known of their production, turnover and physiological structure, very little is understood regarding the three-dimensional arrangement of collagen molecules in naturally occurring fibrils. This knowledge may provide insight into key biological processes such as fibrillo-genesis and tissue remodeling and into diseases such as heart disease and cancer. Here we present a crystallographic determination of the collagen type I supermolecular structure, where the molecular conformation of each collagen segment found within the naturally occurring crystallographic unit cell has been defined (P1, a approximately 40.0 A, b approximately 27.0 A, c approximately 678 A, alpha approximately 89.2 degrees , beta approximately 94.6 degrees , gamma approximately 105.6 degrees ; reflections: 414, overlapping, 232, and nonoverlapping, 182; resolution, 5.16 A axial and 11.1 A equatorial). This structure shows that the molecular packing topology of the collagen molecule is such that packing neighbors are arranged to form a supertwisted (discontinuous) right-handed microfibril that interdigitates with neighboring microfibrils. This interdigitation establishes the crystallographic superlattice, which is formed of quasihexagonally packed collagen molecules. In addition, the molecular packing structure of collagen shown here provides information concerning the potential modes of action of two prominent molecules involved in human health and disease: decorin and the Matrix Metallo-Proteinase (MMP) collagenase.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
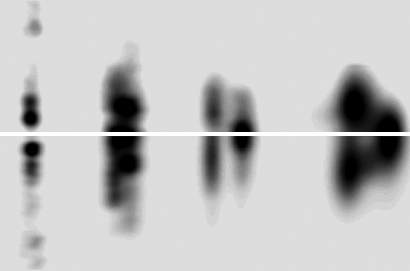
Comment in
-
Re: Microfibrillar structure of type I collagen in situ.Acta Crystallogr D Biol Crystallogr. 2009 Sep;65(Pt 9):1007-8; author reply 1009-10. doi: 10.1107/S0907444909023051. Epub 2009 Aug 14. Acta Crystallogr D Biol Crystallogr. 2009. PMID: 19690380 No abstract available.
References
-
- Hulmes D. J., Miller A. Nature. 1979;282:878–880. - PubMed
-
- North A. C., Cowan P. M., Randall J. T. Nature. 1954;174:1142–1143. - PubMed
-
- Smith J. W. Nature. 1968;219:157–158. - PubMed
-
- Miller A. Biochemistry of Collagen. New York: Plenum; 1976.
-
- Bornstein P., Traub W. The Proteins. London: Academic; 1979.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
