

Unequal sister chromatid and homolog recombination at a tandem duplication of the A1 locus in maize
- PMID: 16751673
- PMCID: PMC1569709
- DOI: 10.1534/genetics.105.052712
Unequal sister chromatid and homolog recombination at a tandem duplication of the A1 locus in maize
Abstract
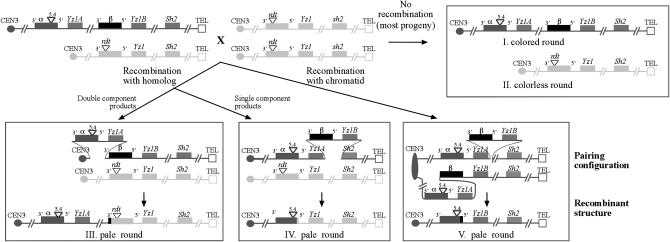
Tandemly arrayed duplicate genes are prevalent. The maize A1-b haplotype is a tandem duplication that consists of the components, alpha and beta. The rate of meiotic unequal recombination at A1-b is ninefold higher when a homolog is present than when it is absent (i.e., hemizygote). When a sequence heterologous homolog is available, 94% of recombinants (264/281) are generated via recombination with the homolog rather than with the sister chromatid. In addition, 83% (220/264) of homolog recombination events involved alpha rather than beta. These results indicate that: (1) the homolog is the preferred template for unequal recombination and (2) pairing of the duplicated segments with the homolog does not occur randomly but instead favors a particular configuration. The choice of recombination template (i.e., homolog vs. sister chromatid) affects the distribution of recombination breakpoints within a1. Rates of unequal recombination at A1-b are similar to the rate of recombination between nonduplicated a1 alleles. Unequal recombination is therefore common and is likely to be responsible for the generation of genetic variability, even within inbred lines.
Figures





References
Publication types
MeSH terms
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
