

The origin and evolution of Archaea: a state of the art
- PMID: 16754611
- PMCID: PMC1578729
- DOI: 10.1098/rstb.2006.1841
The origin and evolution of Archaea: a state of the art
Abstract
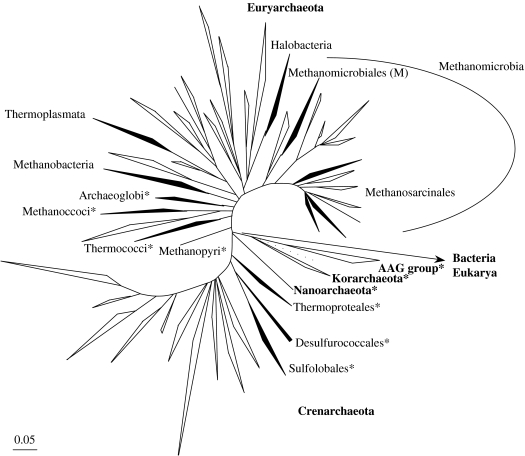
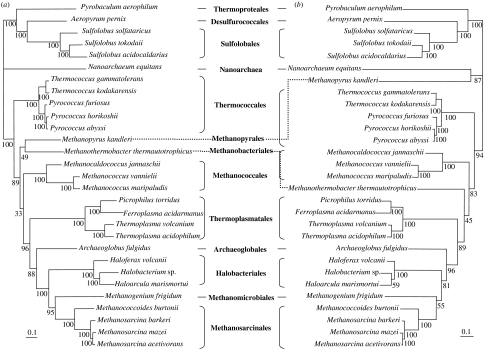
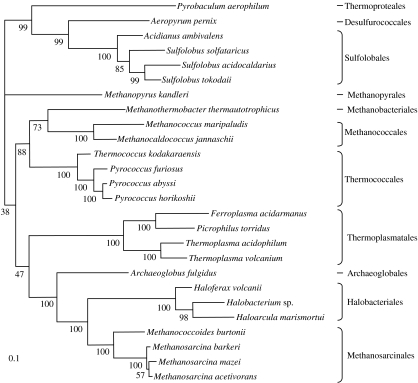
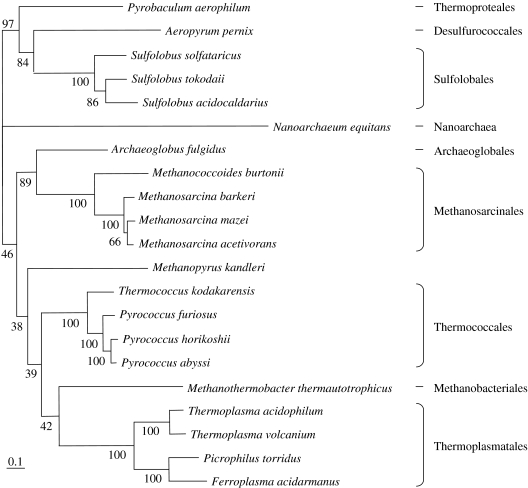
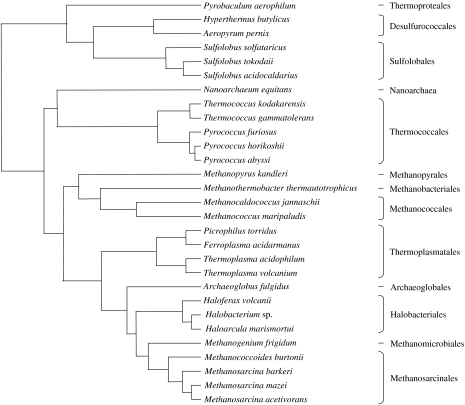
Environmental surveys indicate that the Archaea are diverse and abundant not only in extreme environments, but also in soil, oceans and freshwater, where they may fulfil a key role in the biogeochemical cycles of the planet. Archaea display unique capacities, such as methanogenesis and survival at temperatures higher than 90 degrees C, that make them crucial for understanding the nature of the biota of early Earth. Molecular, genomics and phylogenetics data strengthen Woese's definition of Archaea as a third domain of life in addition to Bacteria and Eukarya. Phylogenomics analyses of the components of different molecular systems are highlighting a core of mainly vertically inherited genes in Archaea. This allows recovering a globally well-resolved picture of archaeal evolution, as opposed to what is observed for Bacteria and Eukarya. This may be due to the fact that no rapid divergence occurred at the emergence of present-day archaeal lineages. This phylogeny supports a hyperthermophilic and non-methanogenic ancestor to present-day archaeal lineages, and a profound divergence between two major phyla, the Crenarchaeota and the Euryarchaeota, that may not have an equivalent in the other two domains of life. Nanoarchaea may not represent a third and ancestral archaeal phylum, but a fast-evolving euryarchaeal lineage. Methanogenesis seems to have appeared only once and early in the evolution of Euryarchaeota. Filling up this picture of archaeal evolution by adding presently uncultivated species, and placing it back in geological time remain two essential goals for the future.
Figures
References
-
- Allmang C, Kufel J, Chanfreau G, Mitchell P, Petfalski E, Tollervey D. Functions of the exosome in rRNA, snoRNA and snRNA synthesis. EMBO J. 1999a;18:5399–5410. 10.1093/emboj/18.19.5399 - DOI - PMC - PubMed
-
- Arrhenius T, Arrhenius G, Paplawsky W. Archean geochemistry of formaldehyde and cyanide and the oligomerization of cyanohydrin. Orig. Life Evol. Biosph. 1994;24:1–17. 10.1007/BF01582036 - DOI - PubMed
-
- Atomi H, Matsumi R, Imanaka T. Reverse gyrase is not a prerequisite for hyperthermophilic life. J. Bacteriol. 2004;186:4829–4833. 10.1128/JB.186.14.4829-4833.2004 - DOI - PMC - PubMed
-
- Bapteste E, Brochier C. On the conceptual difficulties in rooting the tree of life. Trends Microbiol. 2004;12:9–13. 10.1016/j.tim.2003.11.002 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
