

Endoplasmic reticulum aminopeptidase 1 (ERAP1) trims MHC class I-presented peptides in vivo and plays an important role in immunodominance
- PMID: 16754858
- PMCID: PMC1482590
- DOI: 10.1073/pnas.0603095103
Endoplasmic reticulum aminopeptidase 1 (ERAP1) trims MHC class I-presented peptides in vivo and plays an important role in immunodominance
Abstract
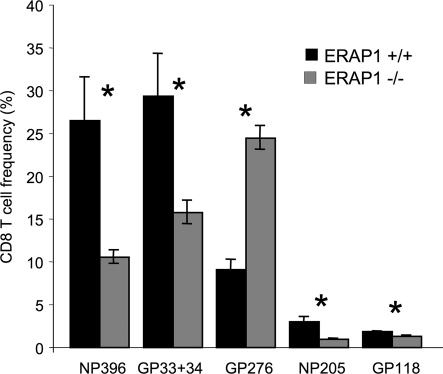
CD8(+) T cells respond to short peptides bound to MHC class I molecules. Although most antigenic proteins contain many sequences that could bind to MHC class I, few of these peptides actually stimulate CD8(+) T cell responses. Moreover, the T cell responses that are generated often follow a very reproducible hierarchy to different peptides for reasons that are poorly understood. We find that the loss of a single enzyme, endoplasmic reticulum aminopeptidase 1 (ERAP1), in the antigen-processing pathway results in a marked shift in the hierarchy of immunodominance in viral infections, even when the responding T cells have the same T cell receptor repertoire. In mice, ERAP1 is the major enzyme that trims precursor peptides in the endoplasmic reticulum and, in this process, can generate or destroy antigenic peptides. Consequently, when ERAP1 is lost, the immune response to some viral peptides is reduced, to others increased, and to yet others unchanged. Therefore, many epitopes must be initially generated as precursors that are normally trimmed by ERAP1 before binding to MHC class I, whereas others are normally degraded by ERAP1 to lengths that are too short to bind to MHC class I. Moreover, peptide trimming and the resulting abundance of peptide-MHC complexes are dominant factors in establishing immunodominance.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
-
- Yewdell J. W., Bennink J. R. Annu. Rev. Immunol. 1999;17:51–88. - PubMed
-
- Busch D. H., Pamer E. G. J. Immunol. 1998;160:4441–4448. - PubMed
-
- Sandberg J. K., Grufman P., Wolpert E. Z., Franksson L., Chambers B. J., Karre K. J. Immunol. 1998;160:3163–3169. - PubMed
-
- Brehm M. A., Pinto A. K., Daniels K. A., Schneck J. P., Welsh R. M., Selin L. K. Nat. Immunol. 2002;3:627–634. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
