

Centrosome-associated RNA in surf clam oocytes
- PMID: 16754862
- PMCID: PMC1482561
- DOI: 10.1073/pnas.0602859103
Centrosome-associated RNA in surf clam oocytes
Abstract
Centrosomes are the major microtubule-organizing center in animal cells. They are composed of a pair of [9(3) + 0] centrioles surrounded by a relatively ill-defined pericentriolar matrix, provide the ciliary centriole-kinetosome (basal body) progenitor, and organize the assembly of microtubules into the mitotic spindle during cell division. Despite >100 years of microscopic observation and their obvious significance, our understanding of centrosome composition, dynamic organization, and mechanism of action is limited when compared with that of other cellular organelles. Centrosomes duplicate only once per cell cycle to ensure development of a normal bipolar spindle. The initial event in centrosome duplication is centriole replication, which is generative, semiconservative, and independent of the nucleus. Such observations led to the proposal that centrosomes contain their own complement of nucleic acids, possibly representative of an organellar genome comparable with those described for mitochondria and chloroplasts. The consensus in the field is that centrosomes lack DNA but may contain RNA. We isolated centrosomes from oocytes of the surf clam, Spisula solidissima, and purified from them a unique set of RNAs. We show here by biochemical means and subcellular in situ hybridization that the first transcript we analyzed is intimately associated with centrosomes. Sequence analysis reveals that this centrosome-associated RNA encodes a conserved RNA-directed polymerase domain. The hypothesis that centrosomes contain an intrinsic complement of specific RNAs suggests new opportunities to address the century-old problem of centrosome function, heredity, and evolution.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
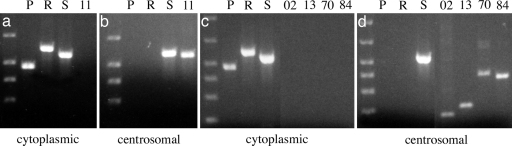
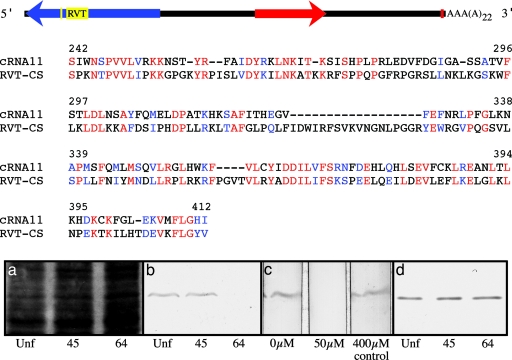
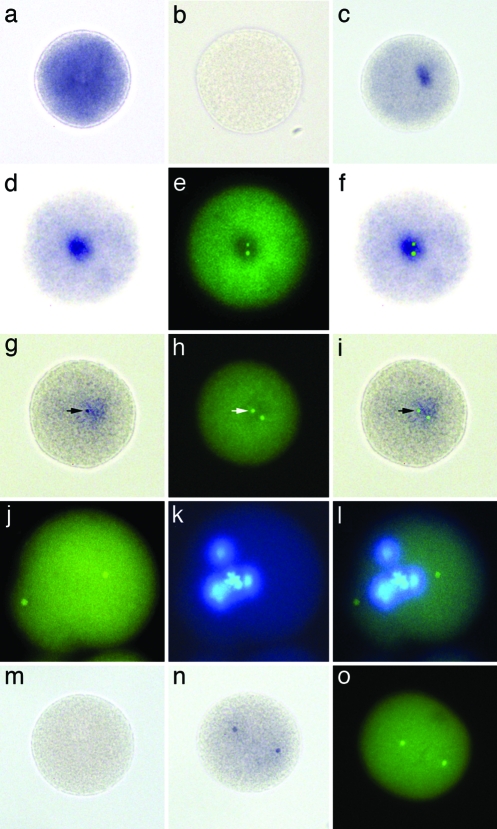
References
-
- Sluder G., Hinchcliffe E. H. Biol. Cell. 1999;91:413–427. - PubMed
-
- Wong C., Stearns T. Nat. Cell Biol. 2003;5:539–544. - PubMed
-
- Chapman M. J., Dolan M. F., Margulis L. Q. Rev. Biol. 2000;75:409–429. - PubMed
-
- Stich H. Chromosoma. 1954;6:199–236. - PubMed
-
- Mazia D. Symp. Soc. Exp. Biol. 1955;9:335–357.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
