

Mutation of the atrophin2 gene in the zebrafish disrupts signaling by fibroblast growth factor during development of the inner ear
- PMID: 16754885
- PMCID: PMC1474007
- DOI: 10.1073/pnas.0603453103
Mutation of the atrophin2 gene in the zebrafish disrupts signaling by fibroblast growth factor during development of the inner ear
Abstract
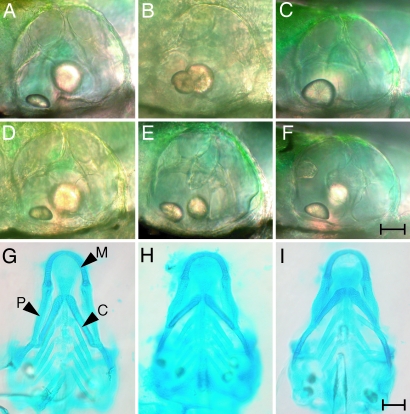
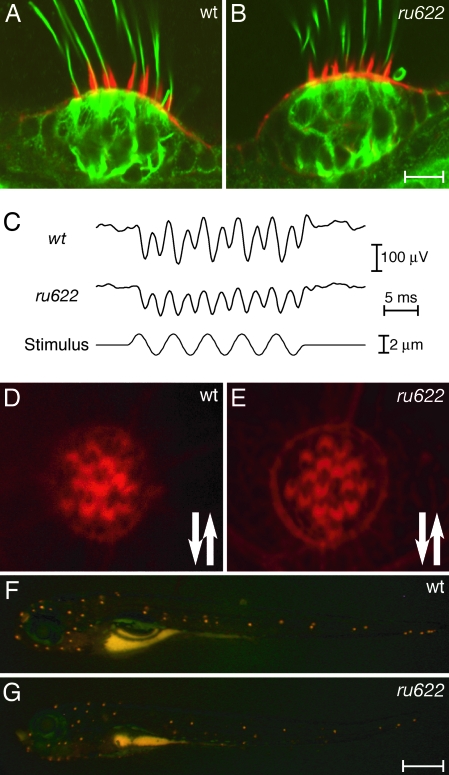
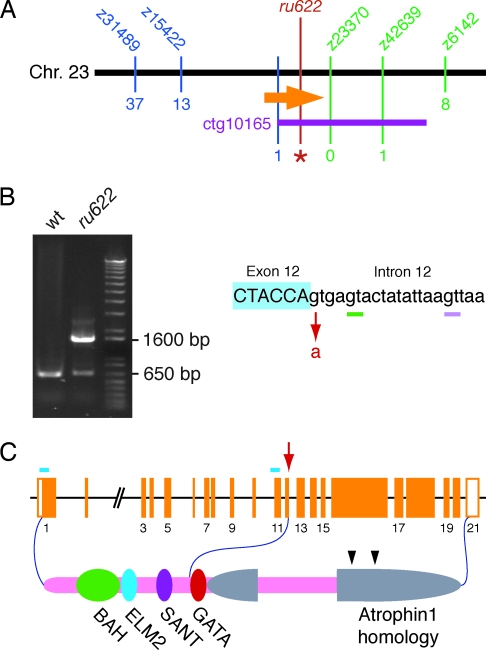
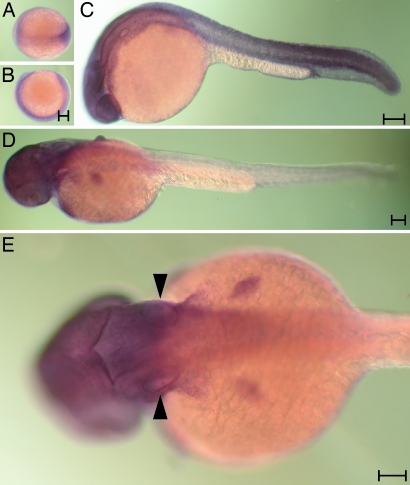
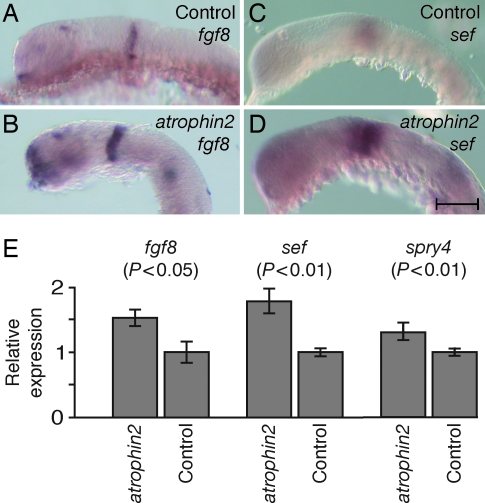
The development of the vertebrate inner ear depends on the precise expression of fibroblast growth factors. In a mutagenesis screen for zebrafish with abnormalities of inner-ear development and behavior, we isolated a mutant line, ru622, whose phenotypic characteristics resembled those of null mutants for the gene encoding fibroblast growth factor 8 (Fgf8): an inconsistent startle response, circular swimming, fused otoliths, and abnormal semicircular canals. Positional cloning disclosed that the mutant gene encodes the transcriptional corepressor Atrophin2. Both the Fgf8 protein and zebrafish "similar expression to fgf genes" protein (Sef), an antagonist of fibroblast growth factors induced by Fgf8 itself, were found to be overexpressed in ru622 mutants. We therefore hypothesized that an excess of Sef eliminates Fgf8 signals and produces an fgf8 null phenotype in ru622 mutants. In support of this idea, we could rescue larvae whose atrophin2 expression had been diminished with morpholinos by reducing the expression of Sef as well. We propose that Atrophin2 plays a role in the feedback regulation of Fgf8 signaling. When mutation of the atrophin2 gene results in the overexpression of both Fgf8 and Sef, the excessive Sef inhibits Fgf8 signaling. The resultant imbalance of Fgf8 and Sef signals then underlies the abnormal aural development observed in ru622.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
-
- Hudspeth A. J. Nature. 1998;341:397–404. - PubMed
-
- Barald K. F., Kelley M. W. Development (Cambridge, U.K.) 2004;131:4119–4130. - PubMed
-
- Haddon C., Lewis J. J. Comp. Neurol. 1996;365:113–128. - PubMed
-
- Whitfield T. T., Riley B. B., Chiang M.-Y., Phillips B. Dev. Dyn. 2002;223:427–458. - PubMed
-
- Haddon C., Lewis J. Development (Cambridge, U.K.) 1991;112:541–550. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
