

Ups1p, a conserved intermembrane space protein, regulates mitochondrial shape and alternative topogenesis of Mgm1p
- PMID: 16754953
- PMCID: PMC2063882
- DOI: 10.1083/jcb.200603092
Ups1p, a conserved intermembrane space protein, regulates mitochondrial shape and alternative topogenesis of Mgm1p
Abstract
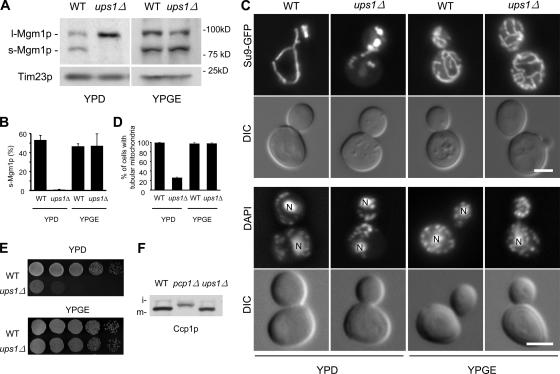
Mgm1p is a conserved dynamin-related GTPase required for fusion, morphology, inheritance, and the genome maintenance of mitochondria in Saccharomyces cerevisiae. Mgm1p undergoes unconventional processing to produce two functional isoforms by alternative topogenesis. Alternative topogenesis involves bifurcate sorting in the inner membrane and intramembrane proteolysis by the rhomboid protease Pcp1p. Here, we identify Ups1p, a novel mitochondrial protein required for the unique processing of Mgm1p and for normal mitochondrial shape. Our results demonstrate that Ups1p regulates the sorting of Mgm1p in the inner membrane. Consistent with its function, Ups1p is peripherally associated with the inner membrane in the intermembrane space. Moreover, the human homologue of Ups1p, PRELI, can fully replace Ups1p in yeast cells. Together, our findings provide a conserved mechanism for the alternative topogenesis of Mgm1p and control of mitochondrial morphology.
Figures




References
-
- Adams, A., D. Gottschling, C. Kaiser, and T. Stearns. 1997. Methods in Yeast Genetics. Cold Spring Harbor Laboratory Press, Plainview, NY. 177 pp.
-
- Alexander, C., M. Votruba, U.E. Pesch, D.L. Thiselton, S. Mayer, A. Moore, M. Rodriguez, U. Kellner, B. Leo-Kottler, G. Auburger, et al. 2000. OPA1, encoding a dynamin-related GTPase, is mutated in autosomal dominant optic atrophy linked to chromosome 3q28. Nat. Genet. 26:211–215. - PubMed
-
- Brachmann, C.B., A. Davies, G.J. Cost, E. Caputo, J. Li, P. Hieter, and J.D. Boeke. 1998. Designer deletion strains derived from Saccharomyces cerevisiae 288C: a useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast. 14:115–132. - PubMed
-
- Chen, H., and D.C. Chan. 2004. Mitochondrial dynamics in mammals. Curr. Top. Dev. Biol. 59:119–144. - PubMed
-
- Delettre, C., G. Lenaers, J.M. Griffoin, N. Gigarel, C. Lorenzo, P. Belenguer, L. Pelloquin, J. Grosgeorge, C. Turc-Carel, E. Perret, et al. 2000. Nuclear gene OPA1, encoding a mitochondrial dynamin-related protein, is mutated in dominant optic atrophy. Nat. Genet. 26:207–210. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
