

Morphological analysis of the hindlimb in apes and humans. I. Muscle architecture
- PMID: 16761973
- PMCID: PMC2100225
- DOI: 10.1111/j.1469-7580.2006.00563.x
Morphological analysis of the hindlimb in apes and humans. I. Muscle architecture
Abstract
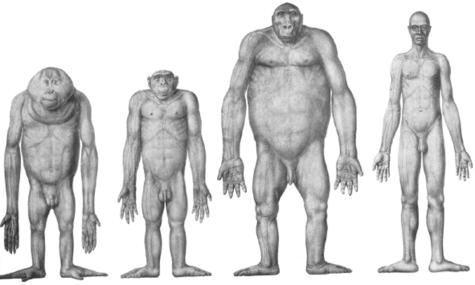
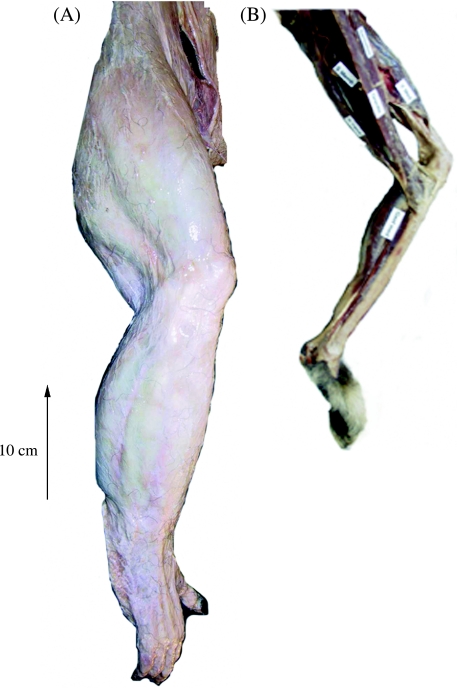
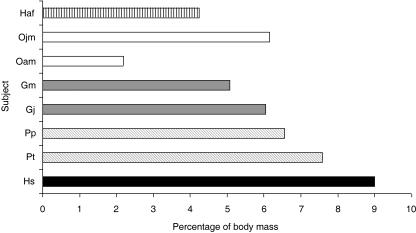
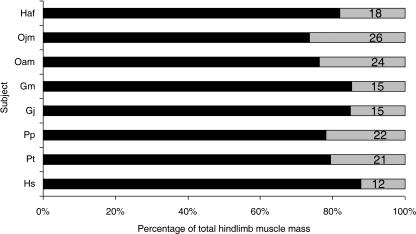
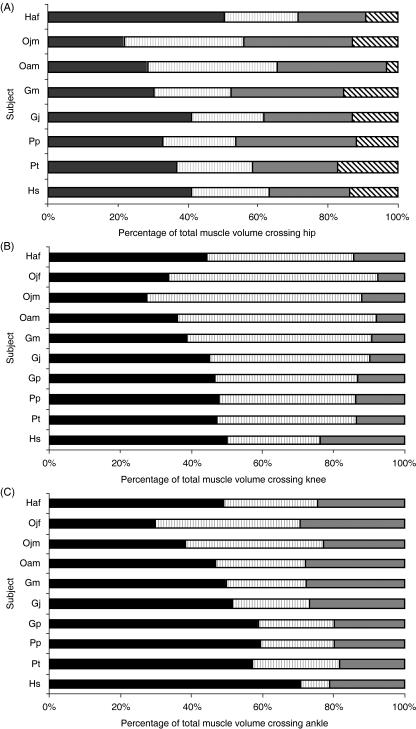
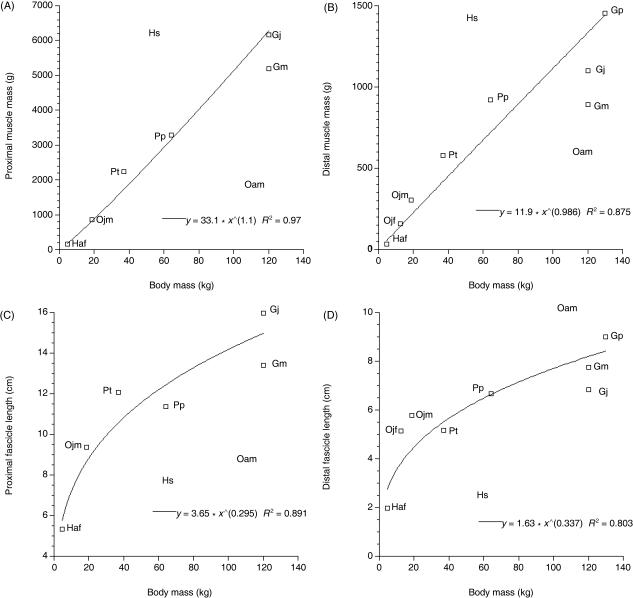
We present quantitative data on the hindlimb musculature of Pan paniscus, Gorilla gorilla gorilla, Gorilla gorilla graueri, Pongo pygmaeus abelii and Hylobates lar and discuss the findings in relation to the locomotor habits of each. Muscle mass and fascicle length data were obtained for all major hindlimb muscles. Physiological cross-sectional area (PCSA) was estimated. Data were normalized assuming geometric similarity to allow for comparison of animals of different size/species. Muscle mass scaled closely to (body mass)(1.0) and fascicle length scaled closely to (body mass)(0.3) in most species. However, human hindlimb muscles were heavy and had short fascicles per unit body mass when compared with non-human apes. Gibbon hindlimb anatomy shared some features with human hindlimbs that were not observed in the non-human great apes: limb circumferences tapered from proximal-to-distal, fascicle lengths were short per unit body mass and tendons were relatively long. Non-human great ape hindlimb muscles were, by contrast, characterized by long fascicles arranged in parallel, with little/no tendon of insertion. Such an arrangement of muscle architecture would be useful for locomotion in a three dimensionally complex arboreal environment.
Figures
References
-
- Aerts P. Vertical jumping in Galago senegalensis: the quest for a hidden power amplifier. Phil Trans R Soc Lond. 1998;353:1607–1620.
-
- Aerts P, Van Damme R, Van Elsacker L, Duchene V. Spatio-temporal gait characteristics of hind-limb cycles during voluntary bipedal and quadrupedal walking in bonobos (Pan paniscus). Am J Phys Anthropol. 2000;111:503–517. - PubMed
-
- Aiello LC, Wheeler P. The expensive-tissue hypothesis. Curr Anthropol. 1995;36:199–221.
-
- Aiello LC, Wells JCK. Energetics and the evolution of the genus Homo. Annu Rev Anthropol. 2002;31:323–338.
-
- Alexander RM, Vernon A. The dimensions of the knee and ankle muscles and the forces they exert. J Hum Mov Stud. 1975;1:115–123.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
