

Antagonism or synergism. Role of tyrosine phosphatases SHP-1 and SHP-2 in growth factor signaling
- PMID: 16762922
- PMCID: PMC2377019
- DOI: 10.1074/jbc.M605018200
Antagonism or synergism. Role of tyrosine phosphatases SHP-1 and SHP-2 in growth factor signaling
Abstract
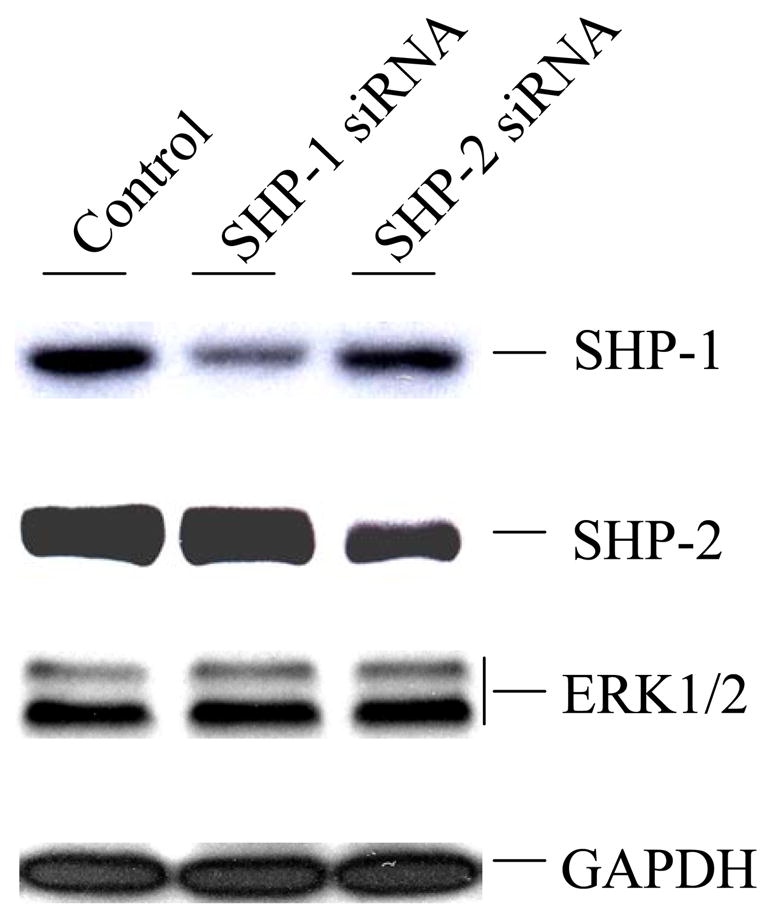
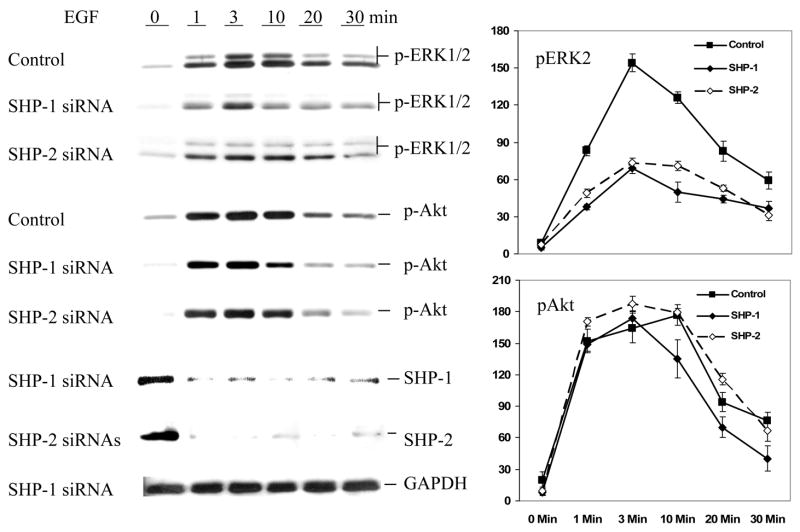
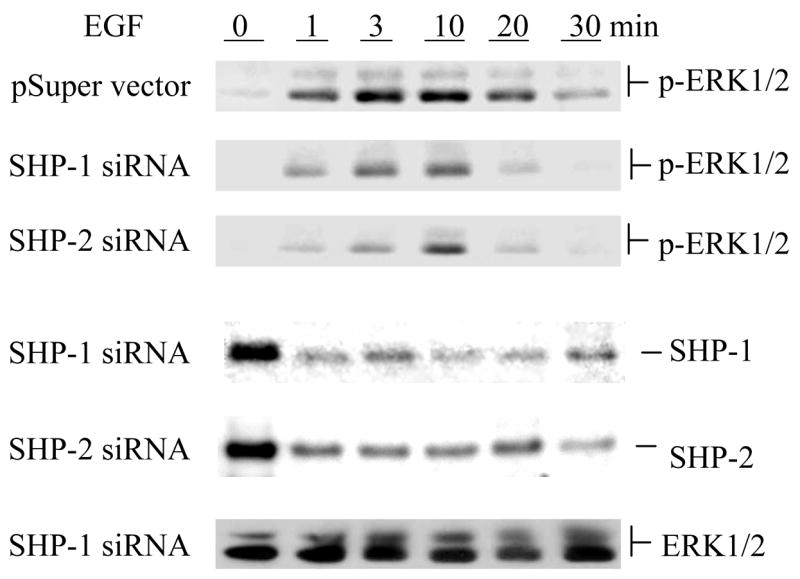
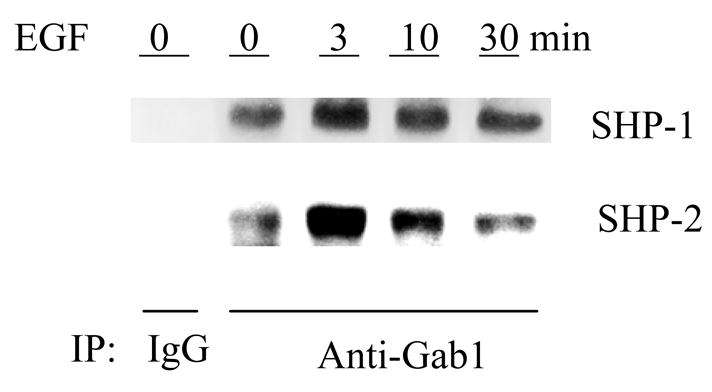
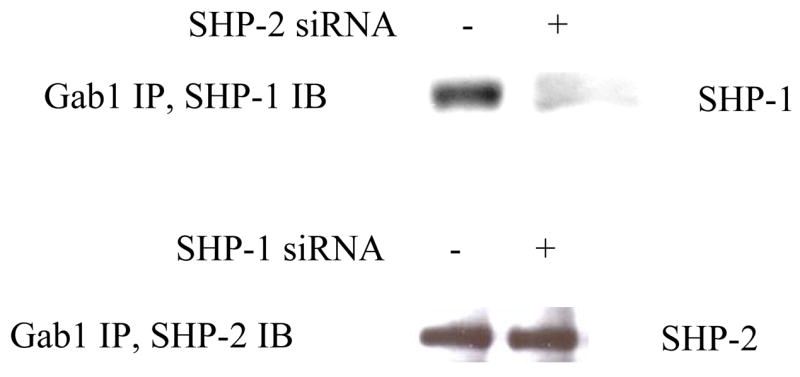
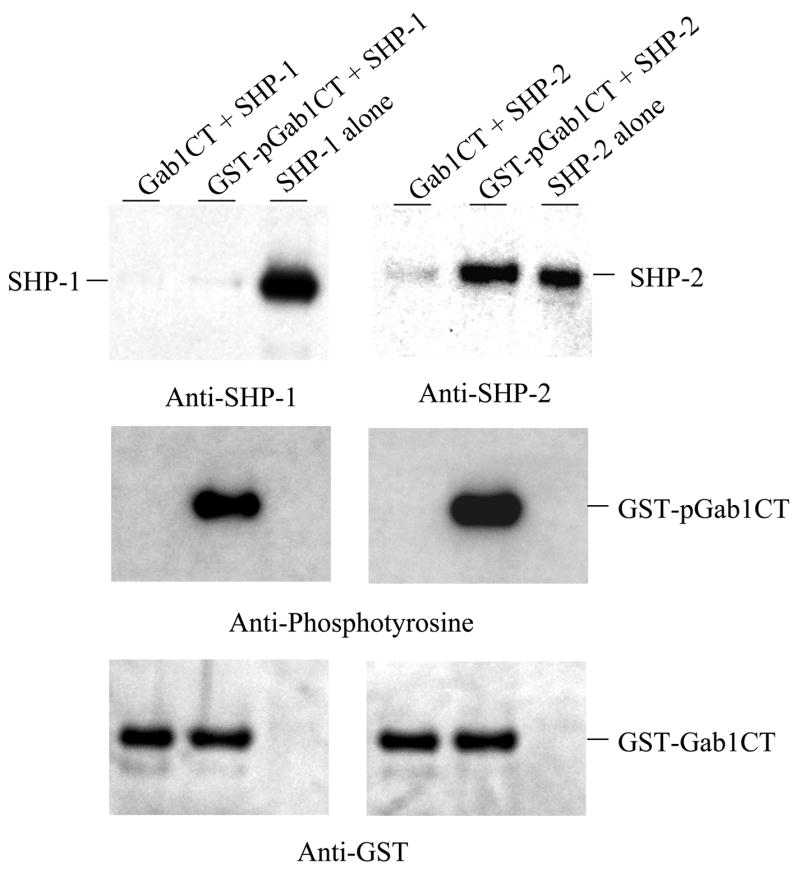
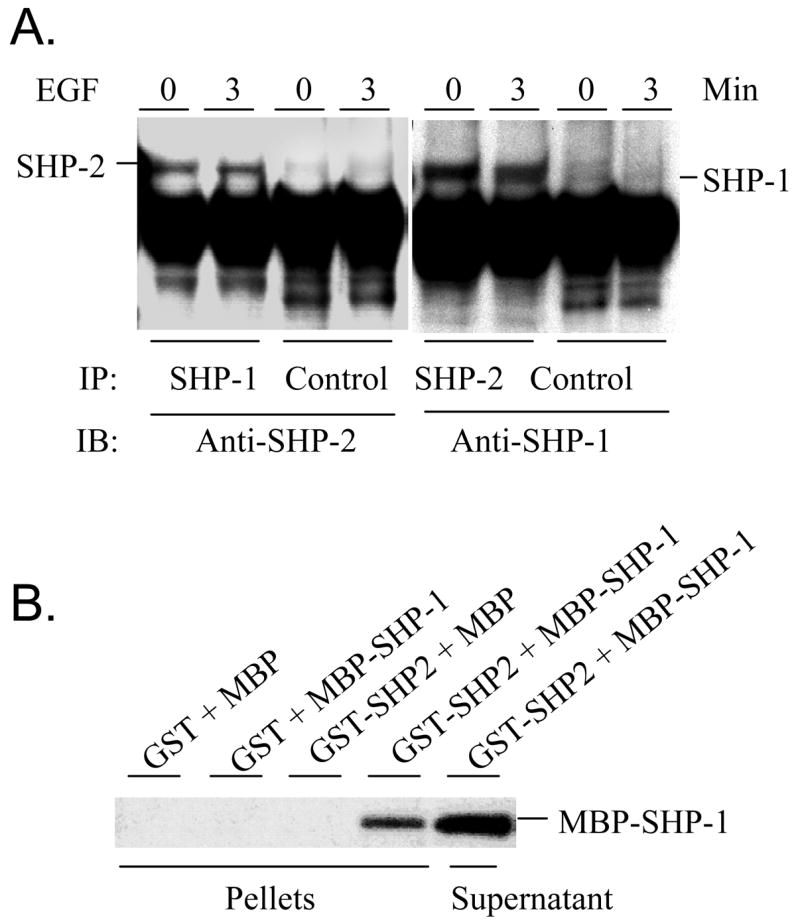
SHP-1 and SHP-2 are two Src homology 2 domain-containing tyrosine phosphatases with major pathological implications in cell growth regulating signaling. They share significant overall sequence identity, but their biological functions are often opposite. SHP-1 is generally considered as a negative signal transducer and SHP-2 as a positive one. However, the precise role of each enzyme in shared signaling pathways is not well defined. In this study, we investigated the interaction of these two enzymes in a single cell system by knocking down their expressions with small interfering RNAs and analyzing the effects on epidermal growth factor signaling. Interestingly, knockdown of either SHP-1 or SHP-2 caused significant reduction in the activation of ERK1/2 but not Akt. Furthermore, SHP-1, SHP-2, and Gab1 formed a signaling complex, and SHP-1 and SHP-2 interact with each other. The interaction of SHP-1 with Gab1 is mediated by SHP-2 because it was abrogated by knockdown of SHP-2, and SHP-2, but not SHP-1, binds directly to tyrosine-phosphorylated Gab1. Together, the data revealed that both SHP-1 and SHP-2 have a positive role in epidermal growth factor-induced ERK1/2 activation and that they act cooperatively rather than antagonistically. The interaction of SHP-1 and SHP-2 may be responsible for previously unexpected novel regulatory mechanism of cell signaling by tyrosine phosphatases.
Figures
Similar articles
-
Dual signaling role of the protein tyrosine phosphatase SHP-2 in regulating expression of acute-phase plasma proteins by interleukin-6 cytokine receptors in hepatic cells.Mol Cell Biol. 1999 Aug;19(8):5326-38. doi: 10.1128/MCB.19.8.5326. Mol Cell Biol. 1999. PMID: 10409724 Free PMC article.
-
Distinct domains in the SHP-2 phosphatase differentially regulate epidermal growth factor receptor/NF-kappaB activation through Gab1 in glioblastoma cells.Mol Cell Biol. 2004 Jan;24(2):823-36. doi: 10.1128/MCB.24.2.823-836.2004. Mol Cell Biol. 2004. PMID: 14701753 Free PMC article.
-
Role of the Grb2-associated binder 1/SHP-2 interaction in cell growth and transformation.Cancer Res. 2004 Mar 15;64(6):2007-15. doi: 10.1158/0008-5472.can-03-2886. Cancer Res. 2004. PMID: 15026337
-
The Src homology 2 domain tyrosine phosphatases SHP-1 and SHP-2: diversified control of cell growth, inflammation, and injury.Histol Histopathol. 2007 Nov;22(11):1251-67. doi: 10.14670/HH-22.1251. Histol Histopathol. 2007. PMID: 17647198 Free PMC article. Review.
-
Immune signalling: SHP-2 docks at multiple ports.Curr Biol. 1999 Feb 25;9(4):R129-32. doi: 10.1016/s0960-9822(99)80080-3. Curr Biol. 1999. PMID: 10074424 Review.
Cited by
-
Arsenite and insulin exhibit opposing effects on epidermal growth factor receptor and keratinocyte proliferative potential.Toxicol Appl Pharmacol. 2007 May 15;221(1):119-28. doi: 10.1016/j.taap.2007.02.003. Epub 2007 Feb 14. Toxicol Appl Pharmacol. 2007. PMID: 17400267 Free PMC article.
-
SHP-2 expression negatively regulates NK cell function.J Immunol. 2009 Dec 1;183(11):7234-43. doi: 10.4049/jimmunol.0900088. Epub 2009 Nov 13. J Immunol. 2009. PMID: 19915046 Free PMC article.
-
PTPN6 expression is epigenetically regulated and influences survival and response to chemotherapy in high-grade gliomas.Tumour Biol. 2014 May;35(5):4479-88. doi: 10.1007/s13277-013-1590-5. Epub 2014 Feb 9. Tumour Biol. 2014. PMID: 24510345
-
Suppression of proximal T cell receptor signaling and lytic function in CD8+ tumor-infiltrating T cells.Cancer Res. 2007 Dec 1;67(23):11447-54. doi: 10.1158/0008-5472.CAN-07-1441. Cancer Res. 2007. PMID: 18056473 Free PMC article.
-
Inferring gene and protein interactions using PubMed citations and consensus Bayesian networks.PLoS One. 2017 Oct 19;12(10):e0186004. doi: 10.1371/journal.pone.0186004. eCollection 2017. PLoS One. 2017. PMID: 29049295 Free PMC article.
References
-
- Neel BG, Gu H, Pao L. Trends Biochem Sci. 2003;28:284–93. - PubMed
-
- Tsui HW, Siminovitch KA, Souza L, Tsui FWL. Nature Genetics. 1993;4:124–129. - PubMed
-
- Shultz LD, Schweitzer PA, Rajan TV, Yi T, Ihle JN, Matthews RJ, Thomas ML, Beier DR. Cell. 1993;73:1445–1454. - PubMed
-
- Wu C, Sun M, Liu L, Zhou GW. Gene. 2003;306:1–12. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
