

IRBIT, an inositol 1,4,5-trisphosphate receptor-binding protein, specifically binds to and activates pancreas-type Na+/HCO3- cotransporter 1 (pNBC1)
- PMID: 16769890
- PMCID: PMC1480443
- DOI: 10.1073/pnas.0602250103
IRBIT, an inositol 1,4,5-trisphosphate receptor-binding protein, specifically binds to and activates pancreas-type Na+/HCO3- cotransporter 1 (pNBC1)
Abstract
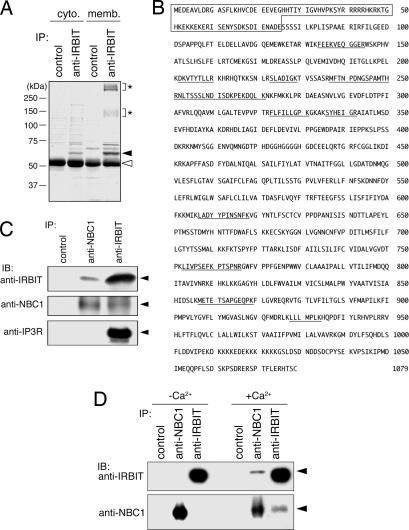
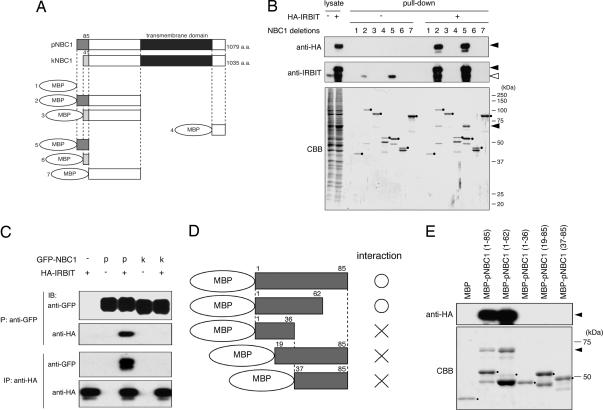
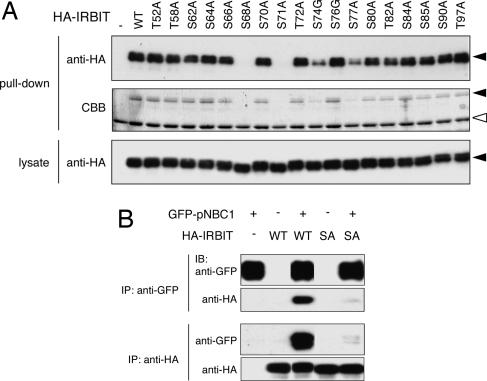
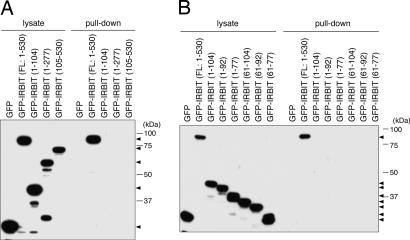
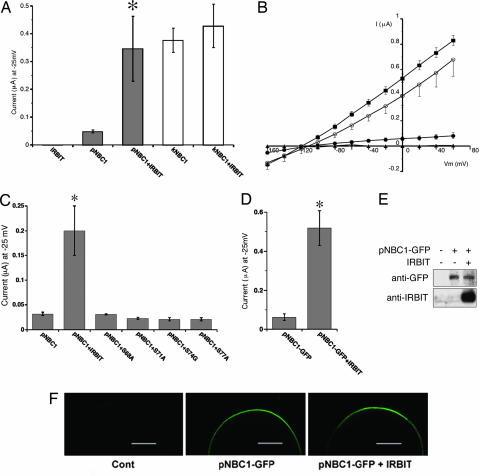
Inositol 1,4,5-trisphosphate (IP(3)) receptors (IP(3)Rs) are IP(3)-gated Ca(2+) channels that are located on intracellular Ca(2+) stores. We previously identified an IP(3)R binding protein, termed IP(3)R binding protein released with IP(3) (IRBIT). Because IRBIT is released from IP(3)R by physiological concentrations of IP(3), we hypothesized that IRBIT is a signaling molecule that is released from IP(3)R and regulates downstream target molecules in response to the production of IP(3). Therefore, in this study, we attempted to identify the target molecules of IRBIT, and we succeeded in identifying Na(+)/HCO(3)(-) cotransporter 1 (NBC1) as an IRBIT binding protein. Of the two major splicing variants of NBC1, pancreas-type NBC1 (pNBC1) and kidney-type NBC1 (kNBC1), IRBIT was found to bind specifically to pNBC1 and not to bind to kNBC1. IRBIT binds to the N-terminal pNBC1-specific domain, and its binding depends on the phosphorylation of multiple serine residues of IRBIT. Also, an electrophysiological analysis in Xenopus oocytes revealed that pNBC1 requires coexpression of IRBIT to manifest substantial activity comparable with that of kNBC1, which displays substantial activity independently of IRBIT. These results strongly suggest that pNBC1 is the target molecule of IRBIT and that IRBIT has an important role in pH regulation through pNBC1. Also, our findings raise the possibility that the regulation through IRBIT enables NBC1 variants to have different physiological roles.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
-
- Berridge M. J. Nature. 1993;361:315–325. - PubMed
-
- Ando H., Mizutani A., Matsu-ura T., Mikoshiba K. J. Biol. Chem. 2003;278:10602–10612. - PubMed
-
- Soleimani M., Burnham C. E. J. Membr. Biol. 2001;183:71–84. - PubMed
-
- Igarashi T., Inatomi J., Sekine T., Cha S. H., Kanai Y., Kunimi M., Tsukamoto K., Satoh H., Shimadzu M., Tozawa F., et al. Nat. Genet. 1999;23:264–266. - PubMed
-
- Igarashi T., Inatomi J., Sekine T., Seki G., Shimadzu M., Tozawa F., Takeshima Y., Takumi T., Takahashi T., Yoshikawa N., et al. J. Am. Soc. Nephrol. 2001;12:713–718. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
