

The PINc domain protein Utp24, a putative nuclease, is required for the early cleavage steps in 18S rRNA maturation
- PMID: 16769905
- PMCID: PMC1480430
- DOI: 10.1073/pnas.0603673103
The PINc domain protein Utp24, a putative nuclease, is required for the early cleavage steps in 18S rRNA maturation
Abstract
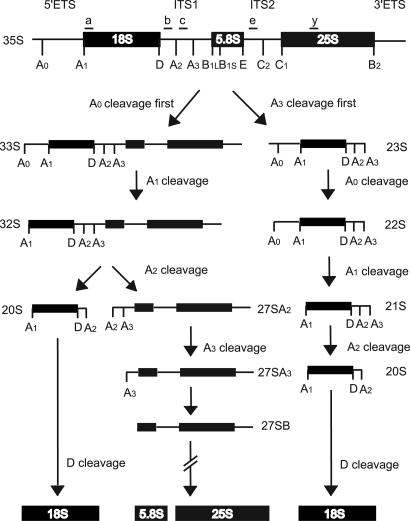
Ribosome biogenesis is a complex process that requires >150 transacting factors, many of which form macromolecular assemblies as big and complex as the ribosome itself. One of those complexes, the SSU processome, is required for pre-18S rRNA maturation. Although many of its components have been identified, the endonucleases that cleave the pre-18S rRNA have remained mysterious. Here we examine the role of four previously uncharacterized PINc domain proteins, which are predicted to function as nucleases, in yeast ribosome biogenesis. We also included Utp23, a protein homologous to the PINc domain protein Utp24, in our analysis. Our results demonstrate that Utp23 and Utp24 are essential nucleolar proteins and previously undescribed components of the SSU processome. In that sense, both Utp23 and Utp24 are required for the first three cleavage steps in 18S rRNA maturation. In addition, single-point mutations in the conserved putative active site of Utp24 but not Utp23 abrogate its function in ribosome biogenesis. Our results suggest that Utp24 might be the elusive endonuclease that cleaves the pre-rRNA at sites A(1) and/or A(2.).
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures





References
-
- Grandi P., Rybin V., Bassler J., Petfalski E., Strauss D., Marzioch M., Schafer T., Kuster B., Tschochner H., Tollervey D., et al. Mol. Cell. 2002;10:105–115. - PubMed
-
- Osheim Y. N., French S. L., Keck K. M., Champion E. A., Spasov K., Dragon F., Baserga S. J., Beyer A. L. Mol. Cell. 2004;16:943–954. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
