

The nucleolar protein Esf2 interacts directly with the DExD/H box RNA helicase, Dbp8, to stimulate ATP hydrolysis
- PMID: 16772403
- PMCID: PMC1483223
- DOI: 10.1093/nar/gkl419
The nucleolar protein Esf2 interacts directly with the DExD/H box RNA helicase, Dbp8, to stimulate ATP hydrolysis
Abstract
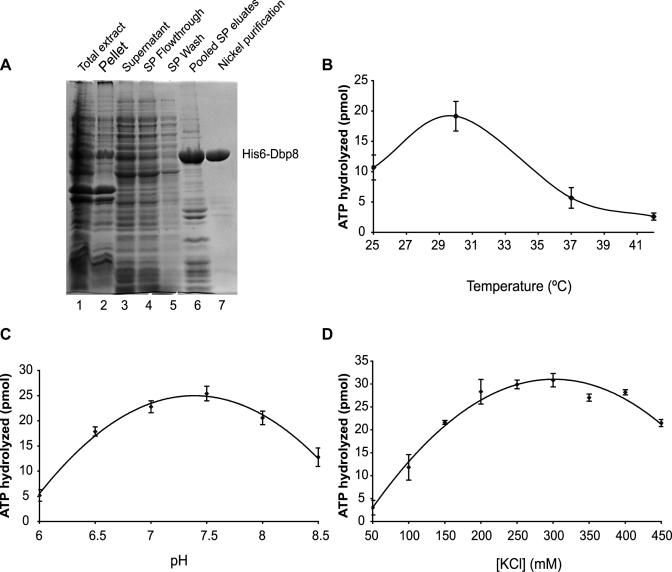
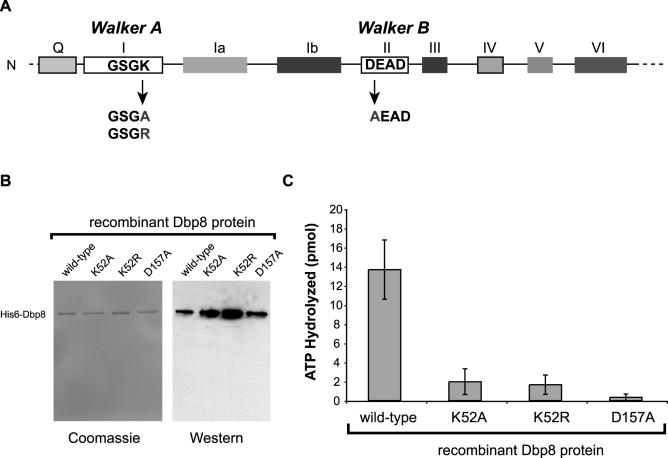
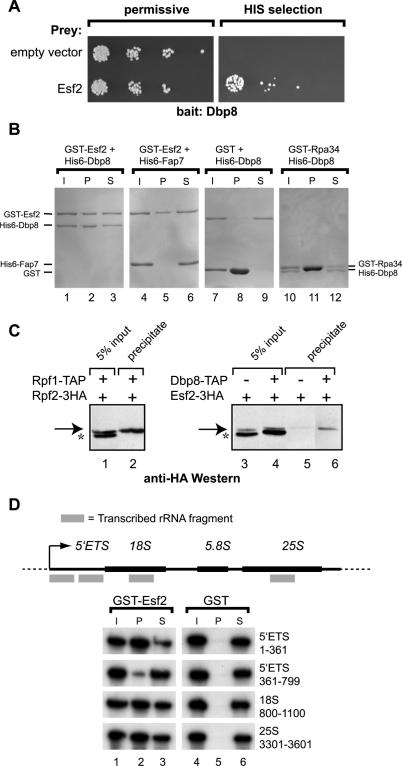
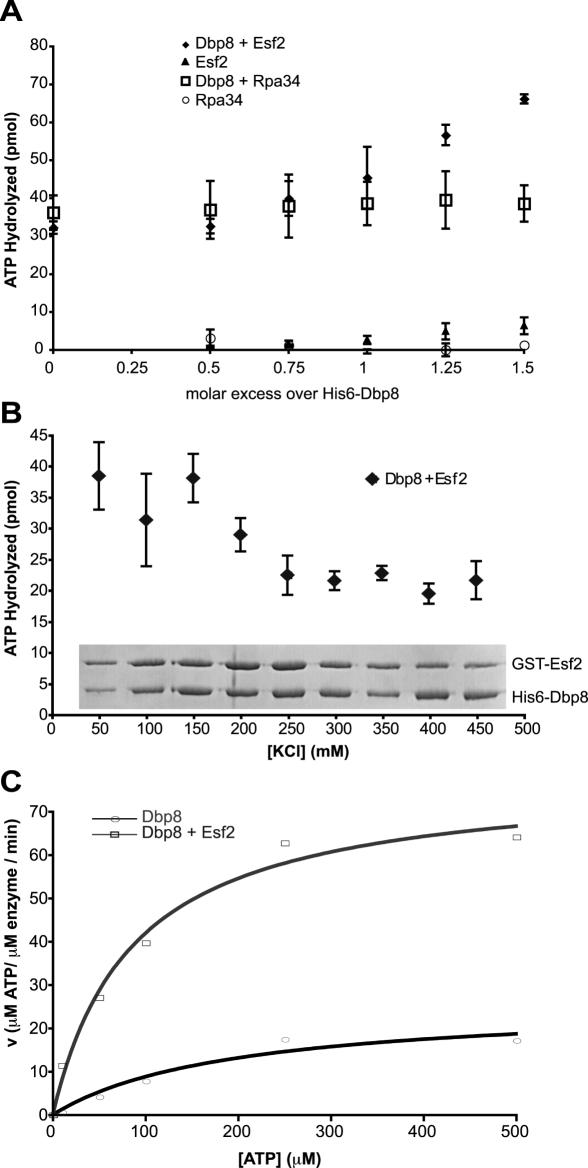
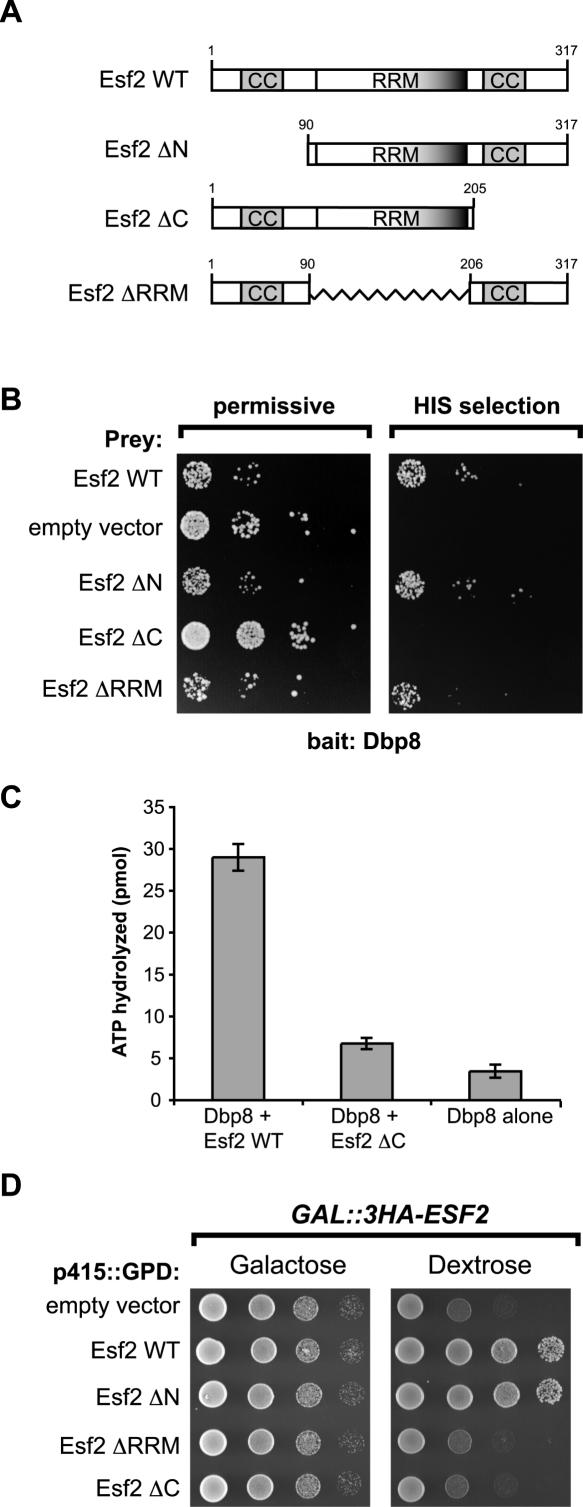
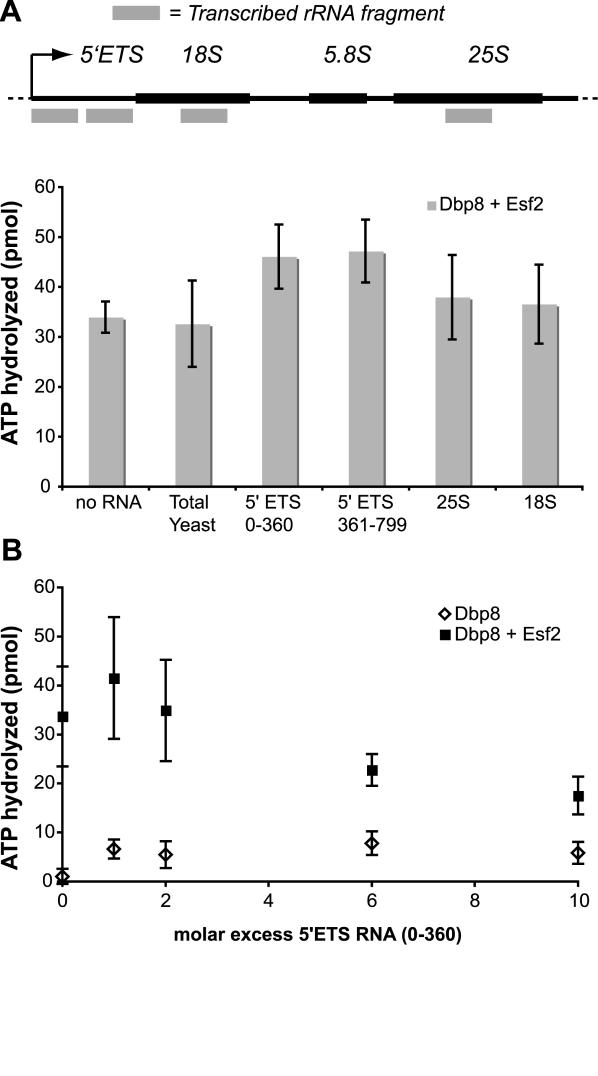
While 18 putative RNA helicases are involved in ribosome biogenesis in Saccharomyces cerevisiae, their enzymatic properties have remained largely biochemically uncharacterized. To better understand their function, we examined the enzymatic properties of Dpb8, a DExD/H box protein previously shown to be required for the synthesis of the 18S rRNA. As expected for an RNA helicase, we demonstrate that recombinant Dbp8 has ATPase activity in vitro, and that this activity is dependent on an intact ATPase domain. Strikingly, we identify Esf2, a nucleolar putative RNA binding protein, as a binding partner for Dbp8, and show that it enhances Dbp8 ATPase activity by decreasing the K(M) for ATP. Thus, we have uncovered Esf2 as the first example of a protein co-factor that has a stimulatory effect on a nucleolar RNA helicase. We show that Esf2 can bind to pre-rRNAs and speculate that it may function to bring Dbp8 to the pre-rRNA, thereby both regulating its enzymatic activity and guiding Dbp8 to its site of action.
Figures
References
-
- Jankowsky E., Gross C.H., Shuman S., Pyle A.M. The DExH protein NPH-II is a processive and directional motor for unwinding RNA. Nature. 2000;403:447–451. - PubMed
-
- Jankowsky E., Gross C.H., Shuman S., Pyle A.M. Active disruption of an RNA-protein interaction by a DExH/D RNA helicase. Science. 2001;291:121–125. - PubMed
-
- Yang Q., Jankowsky E. ATP- and ADP-dependent modulation of RNA unwinding and strand annealing activities by the DEAD-box protein DED1. Biochemistry. 2005;44:13591–13601. - PubMed
-
- Fairman M.E., Maroney P.A., Wang W., Bowers H.A., Gollnick P., Nilsen T.W., Jankowsky E. Protein displacement by DExH/D ‘RNA helicases’ without duplex unwinding. Science. 2004;304:730–734. - PubMed
-
- Rocak S., Linder P. DEAD-box proteins: the driving forces behind RNA metabolism. Nature Rev. Mol. Cell. Biol. 2004;5:232–241. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
