

Testosterone and aggression: Berthold, birds and beyond
- PMID: 16774503
- PMCID: PMC2954190
- DOI: 10.1111/j.1365-2826.2006.01440.x
Testosterone and aggression: Berthold, birds and beyond
Abstract
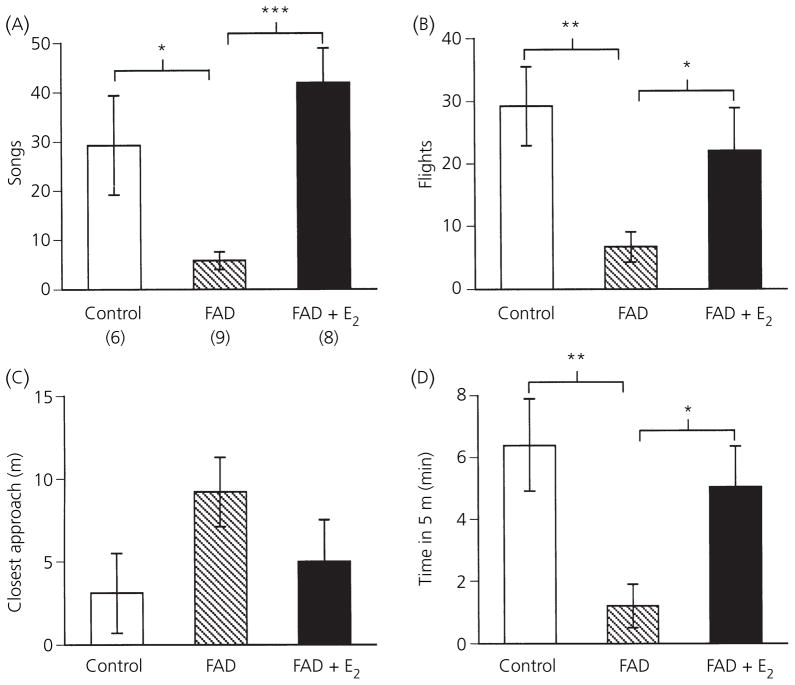
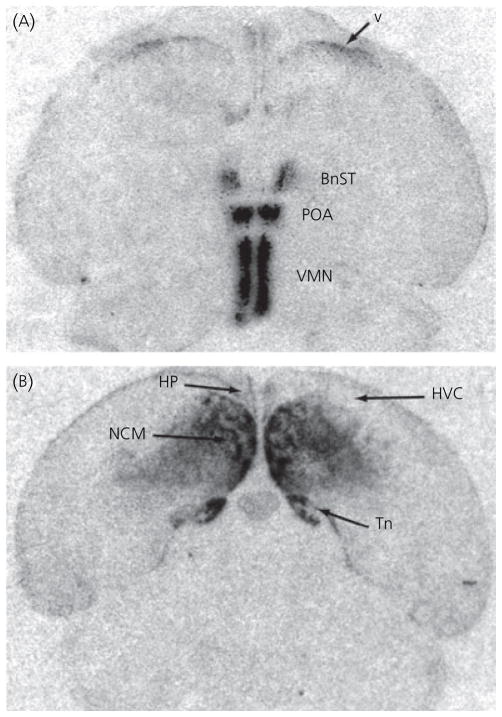
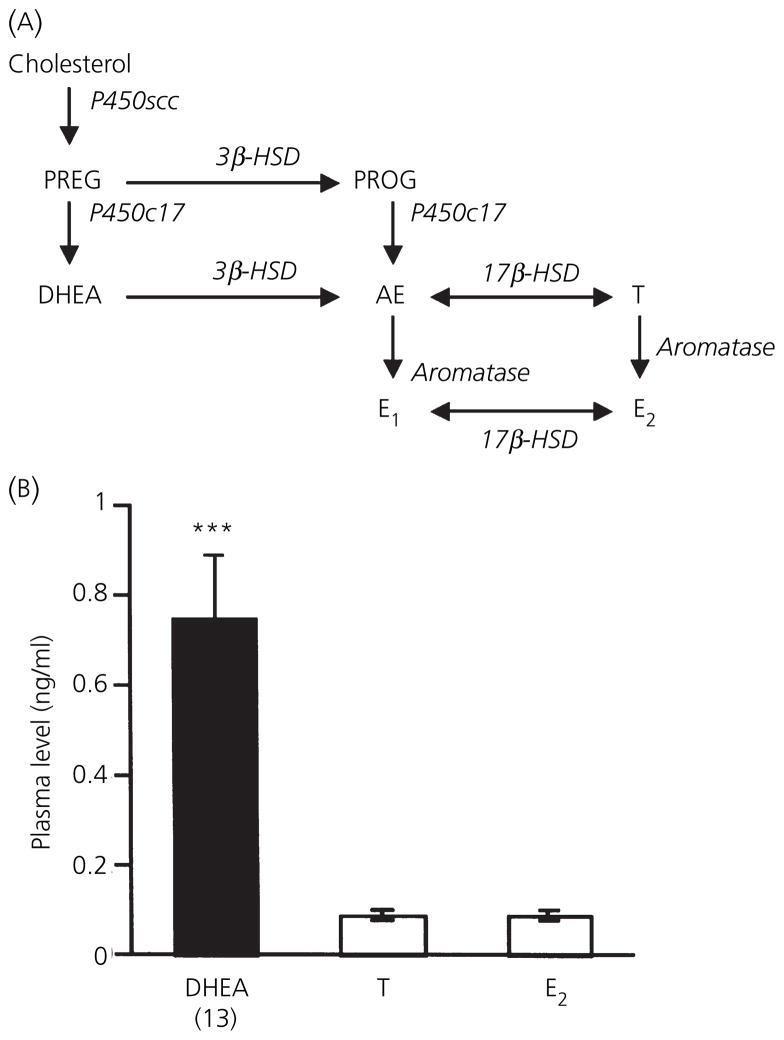
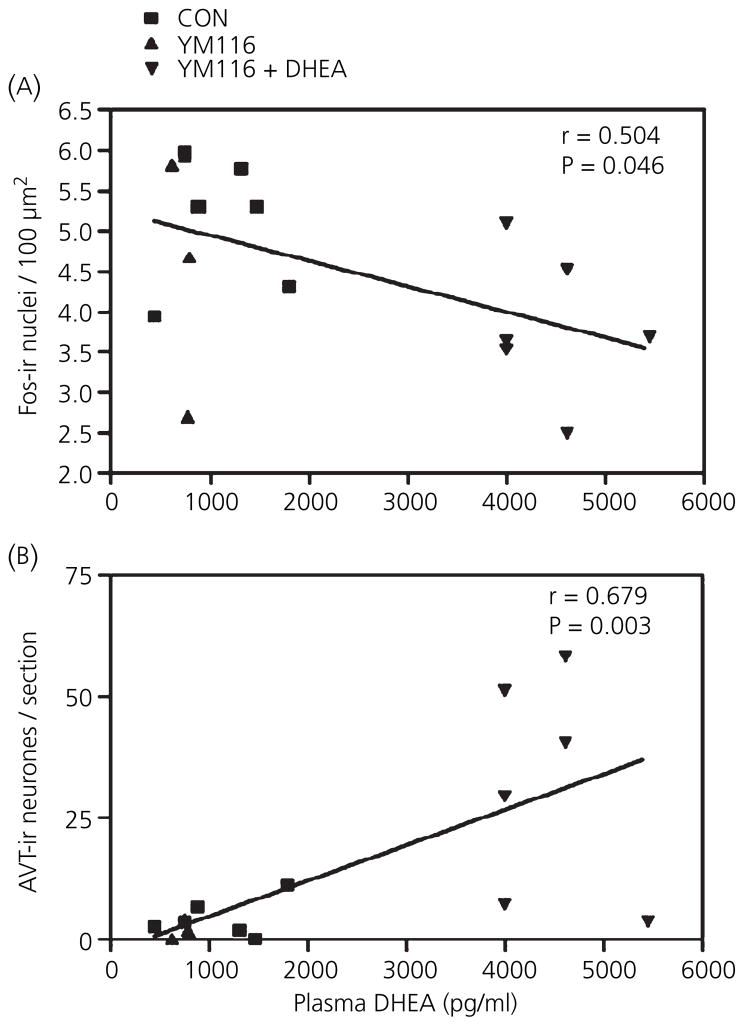
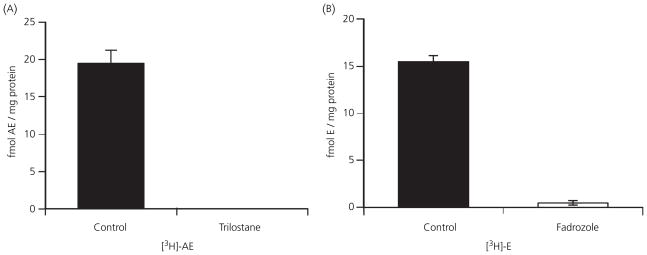
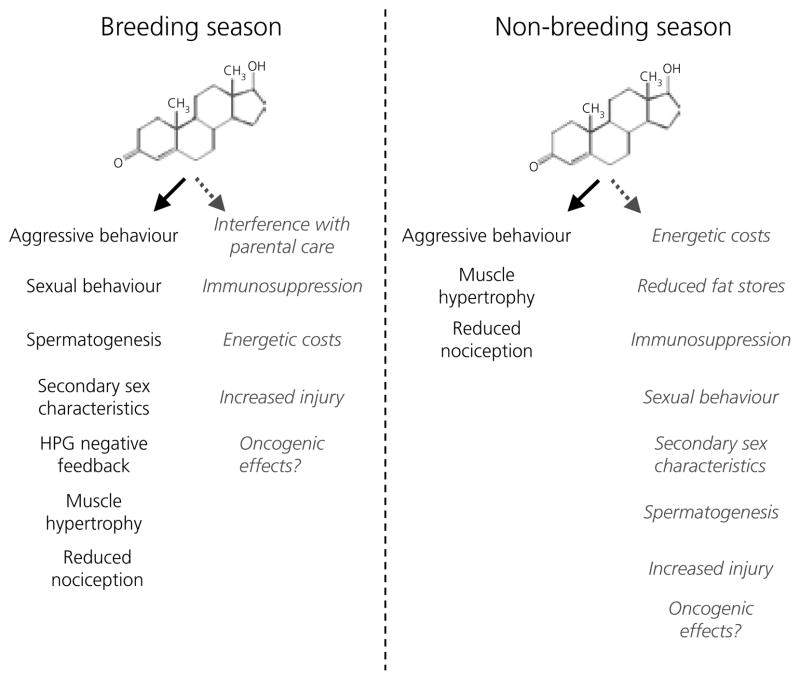
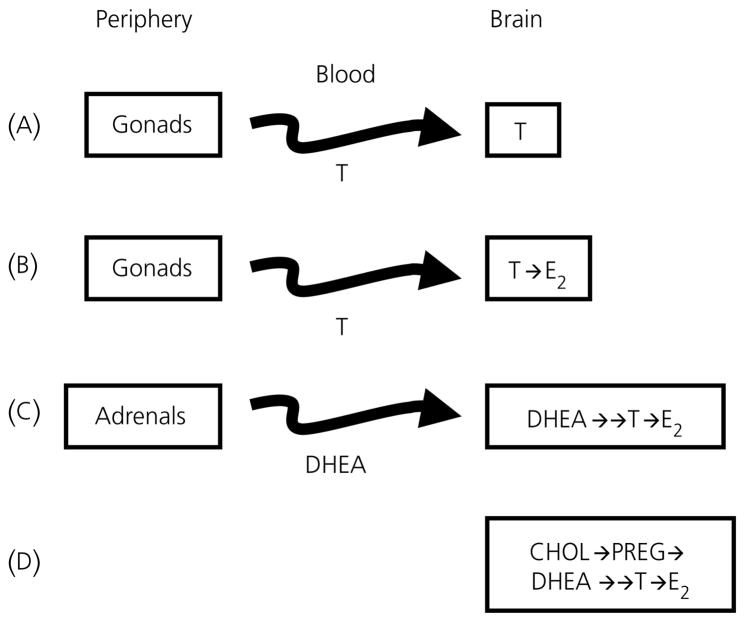
Berthold's classic study of domesticated roosters in 1849 demonstrated that testicular secretions are necessary for the normal expression of aggressive behaviour. Although this conclusion is undoubtedly correct, field studies of wild songbirds have yielded important modifications and limitations of Berthold's original hypothesis. For example, studies of the North American song sparrow (Melospiza melodia) during the breeding season reveal that not only does testosterone increase aggression, but aggressive interactions also increase plasma testosterone levels. Furthermore, in winter, nonbreeding song sparrows have low plasma testosterone levels but are very aggressive, and castration of nonbreeding song sparrows does not decrease aggression. Interestingly, an aromatase inhibitor (fadrozole) does decrease male aggression in the nonbreeding season, and the effects of fadrozole can be rescued with oestradiol. In winter, dehydroepiandrosterone (DHEA) from the periphery can be metabolised within the brain to supply oestradiol to specific neural circuits. Additionally, oestradiol might be synthesised de novo from cholesterol entirely within the brain. These mechanisms may have evolved to avoid the 'costs' of circulating testosterone in the nonbreeding season. Recent studies in tropical birds, hamsters, and humans suggest that these neuroendocrine mechanisms are important for the control of aggression in many vertebrate species.
Figures
References
-
- Wingfield JC, Ball GF, Dufty AM, Hegner RE, Ramenofsky M. Testosterone and aggression in birds. Am Sci. 1987;75:602–608.
-
- Schlinger BA, Callard GV. Aggressive behavior in birds: an experimental model for studies of brain–steroid interactions. Comp Biochem Physiol A. 1990;97:307–316. - PubMed
-
- Harding CF, Follett BK. Hormone changes triggered by aggression in a natural population of blackbirds. Science. 1979;203:918–920. - PubMed
-
- Konishi M, Emlen ST, Ricklefs RE, Wingfield JC. Contributions of bird studies to biology. Science. 1989;246:465–472. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
