

Urinary metabolite profiling reveals CYP1A2-mediated metabolism of NSC686288 (aminoflavone)
- PMID: 16775196
- PMCID: PMC1551906
- DOI: 10.1124/jpet.106.105213
Urinary metabolite profiling reveals CYP1A2-mediated metabolism of NSC686288 (aminoflavone)
Abstract
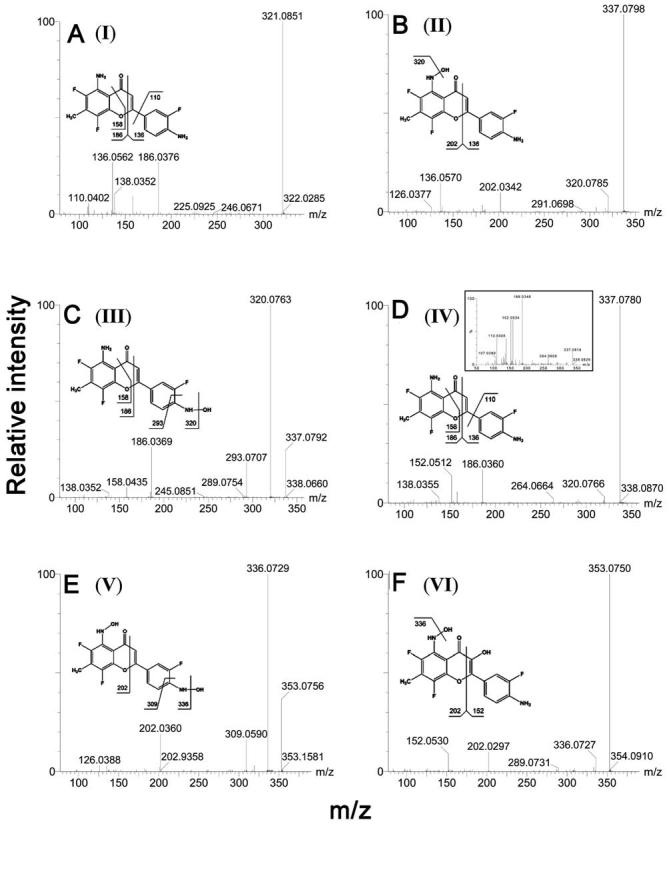
NSC686288 [aminoflavone (AF)], a candidate chemotherapeutic agent, possesses a unique antiproliferative profile against tumor cells. Metabolic bioactivation of AF by drug-metabolizing enzymes, especially CYP1A monooxygenases, has been implicated as an underlying mechanism for its selective cytotoxicity in several cell culture-based studies. However, in vivo metabolism of AF has not been investigated in detail. In this study, the structural identities of 13 AF metabolites (12 of which are novel) in mouse urine or from microsomal incubations, including three monohydroxy-AFs, two dihydroxy-AFs and their sulfate and glucuronide conjugates, as well as one N-glucuronide, were determined by accurate mass measurements and liquid chromatography-tandem mass spectrometry fragmentation patterns, and a comprehensive map of the AF metabolic pathways was constructed. Significant differences between wild-type and Cyp1a2-null mice, within the relative composition of urinary metabolites of AF, demonstrated that CYP1A2-mediated regioselective oxidation was a major contributor to the metabolism of AF. Comparisons between wild-type and CYP1A2-humanized mice further revealed interspecies differences in CYP1A2-mediated catalytic activity. Incubation of AF with liver microsomes from all three mouse lines and with pooled human liver microsomes confirmed the observations from urinary metabolite profiling. Results from enzyme kinetic analysis further indicated that in addition to CYP1A P450s, CYP2C P450s may also play some role in the metabolism of AF.
Figures







References
-
- Akama T, Ishida H, Kimura U, Gomi K, Saito H. Structure-activity relationships of the 7-substituents of 5,4′-diamino-6,8,3′-trifluoroflavone, a potent antitumor agent. J Med Chem. 1998;41:2056–2067. - PubMed
-
- Akama T, Ishida H, Shida Y, Kimura U, Gomi K, Saito H, Fuse E, Kobayashi S, Yoda N, Kasai M. Design and synthesis of potent antitumor 5,4′-diaminoflavone derivatives based on metabolic considerations. J Med Chem. 1997;40:1894–1900. - PubMed
-
- Akama T, Shida Y, Sugaya T, Ishida H, Gomi K, Kasai M. Novel 5-aminoflavone derivatives as specific antitumor agents in breast cancer. J Med Chem. 1996;39:3461–3469. - PubMed
-
- Buters JT, Tang BK, Pineau T, Gelboin HV, Kimura S, Gonzalez FJ. Role of CYP1A2 in caffeine pharmacokinetics and metabolism: studies using mice deficient in CYP1A2. Pharmacogenetics. 1996;6:291–296. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
