

Characterization of the RNA chaperone activity of hantavirus nucleocapsid protein
- PMID: 16775315
- PMCID: PMC1488978
- DOI: 10.1128/JVI.00147-06
Characterization of the RNA chaperone activity of hantavirus nucleocapsid protein
Abstract
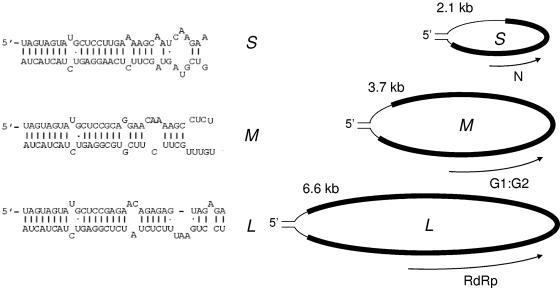
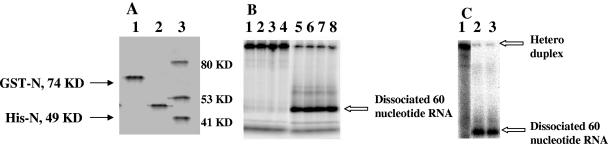
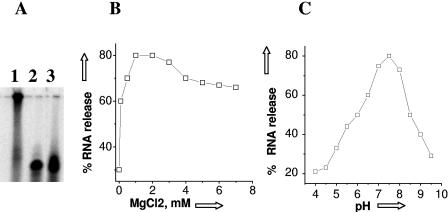
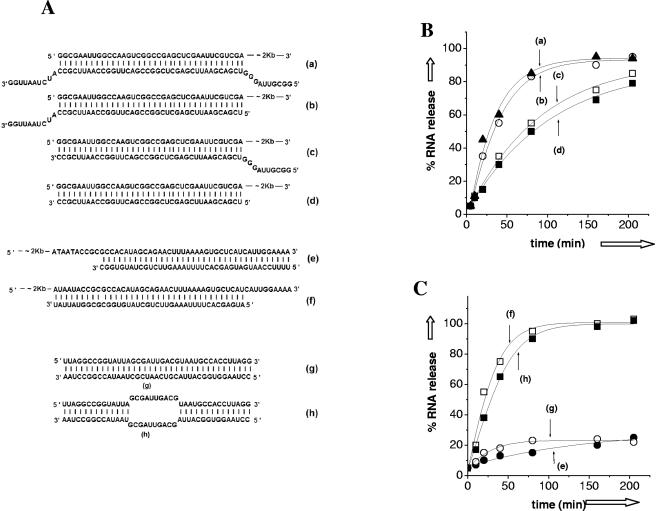
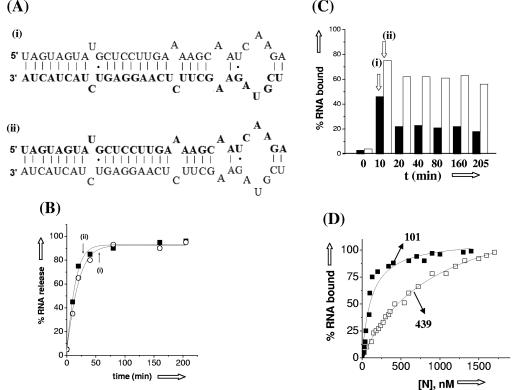
Hantaviruses are tripartite negative-sense RNA viruses and members of the Bunyaviridae family. The nucleocapsid (N) protein, encoded by the smallest of the three genome segments (S), has nonspecific RNA chaperone activity. This activity results in transient dissociation of misfolded RNA structures, may be required for facilitating correct higher-order RNA structure, and may function in viral genome replication. We carried out a series of experiments to further characterize the ability of N to dissociate RNA duplexes. As might be expected, N dissociated RNA duplexes but not DNA duplexes or RNA-DNA heteroduplexes. The RNA-destabilizing activity of N is ATP independent, has a pH optimum of 7.5, and has an Mg(2+) concentration optimum of 1 to 2 mM. N protein is unable to unwind the RNA duplexes that are completely double stranded. However, in the presence of an adjoining single-stranded region, helix unwinding takes place in the 3'-to-5' direction through an unknown mechanism. The N protein trimer specifically recognizes and unwinds the terminal panhandle structure in the viral RNA and remains associated with unwound 5' terminus. We suggest that hantaviral nucleocapsid protein has an active role in hantaviral replication by working cooperatively with viral RNA polymerase. After specific recognition of the panhandle structure by N protein, the unwound 5' terminus likely remains transiently bound to N protein, creating an opportunity for the viral polymerase to initiate transcription at the accessible 3' terminus.
Figures
Similar articles
-
The bunyavirus nucleocapsid protein is an RNA chaperone: possible roles in viral RNA panhandle formation and genome replication.RNA. 2006 Feb;12(2):272-82. doi: 10.1261/rna.2101906. RNA. 2006. PMID: 16428606 Free PMC article.
-
The hantavirus nucleocapsid protein recognizes specific features of the viral RNA panhandle and is altered in conformation upon RNA binding.J Virol. 2005 Feb;79(3):1824-35. doi: 10.1128/JVI.79.3.1824-1835.2005. J Virol. 2005. PMID: 15650206 Free PMC article.
-
Trimeric hantavirus nucleocapsid protein binds specifically to the viral RNA panhandle.J Virol. 2004 Aug;78(15):8281-8. doi: 10.1128/JVI.78.15.8281-8288.2004. J Virol. 2004. PMID: 15254200 Free PMC article.
-
The nucleocapsid of vesicular stomatitis virus.Sci China Life Sci. 2012 Apr;55(4):291-300. doi: 10.1007/s11427-012-4307-x. Epub 2012 May 9. Sci China Life Sci. 2012. PMID: 22566085 Review.
-
Nucleic acid chaperone activity of HIV-1 nucleocapsid protein: critical role in reverse transcription and molecular mechanism.Prog Nucleic Acid Res Mol Biol. 2005;80:217-86. doi: 10.1016/S0079-6603(05)80006-6. Prog Nucleic Acid Res Mol Biol. 2005. PMID: 16164976 Review. No abstract available.
Cited by
-
Comparative nucleic acid chaperone properties of the nucleocapsid protein NCp7 and Tat protein of HIV-1.Virus Res. 2012 Nov;169(2):349-60. doi: 10.1016/j.virusres.2012.06.021. Epub 2012 Jun 26. Virus Res. 2012. PMID: 22743066 Free PMC article. Review.
-
Pseudotyped Viruses for Orthohantavirus.Adv Exp Med Biol. 2023;1407:229-252. doi: 10.1007/978-981-99-0113-5_12. Adv Exp Med Biol. 2023. PMID: 36920700
-
Interaction of hantavirus nucleocapsid protein with ribosomal protein S19.J Virol. 2010 Dec;84(23):12450-3. doi: 10.1128/JVI.01388-10. Epub 2010 Sep 15. J Virol. 2010. PMID: 20844026 Free PMC article.
-
The identification and characterization of nucleic acid chaperone activity of human enterovirus 71 nonstructural protein 3AB.Virology. 2014 Sep;464-465:353-364. doi: 10.1016/j.virol.2014.07.037. Epub 2014 Aug 9. Virology. 2014. PMID: 25113906 Free PMC article.
-
Crimean-Congo hemorrhagic fever virus nucleocapsid protein has dual RNA binding modes.PLoS One. 2017 Sep 18;12(9):e0184935. doi: 10.1371/journal.pone.0184935. eCollection 2017. PLoS One. 2017. PMID: 28922369 Free PMC article.
References
-
- Alfadhli, A., E. Steel, L. Finlay, H. P. Bachinger, and E. Barklis. 2002. Hantavirus nucleocapsid protein coiled-coil domains. J. Biol. Chem. 277:27103-27108. - PubMed
-
- Blakqori, G., G. Kochs, O. Haller, and F. Weber. 2003. Functional L polymerase of La Crosse virus allows in vivo reconstitution of recombinant nucleocapsids. J. Gen. Virol. 84:1207-1214. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
