

Mycovirus cryphonectria hypovirus 1 elements cofractionate with trans-Golgi network membranes of the fungal host Cryphonectria parasitica
- PMID: 16775345
- PMCID: PMC1488963
- DOI: 10.1128/JVI.02519-05
Mycovirus cryphonectria hypovirus 1 elements cofractionate with trans-Golgi network membranes of the fungal host Cryphonectria parasitica
Abstract
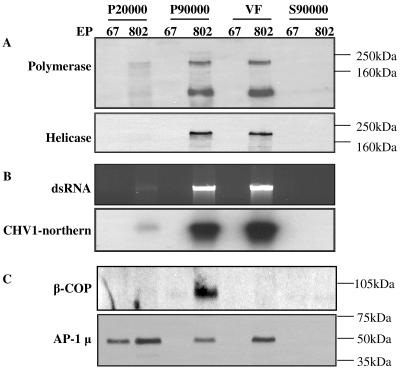
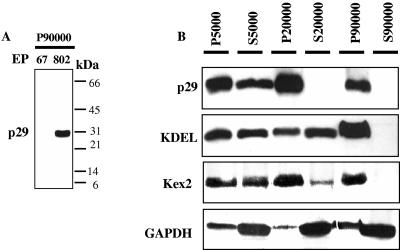
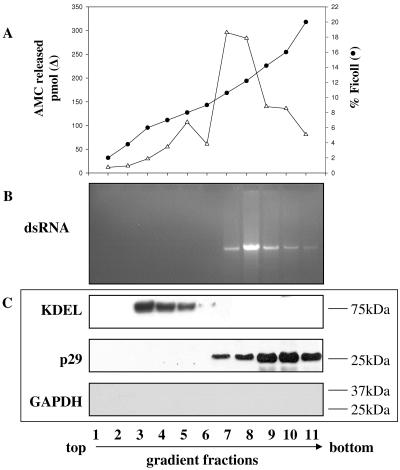
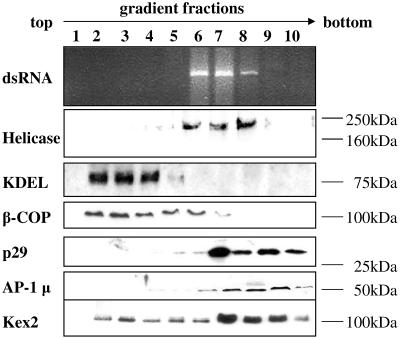
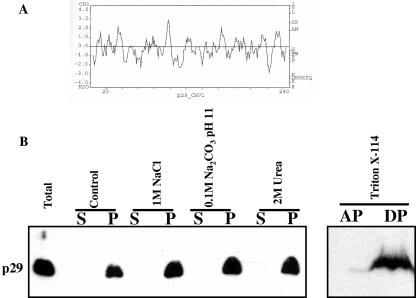
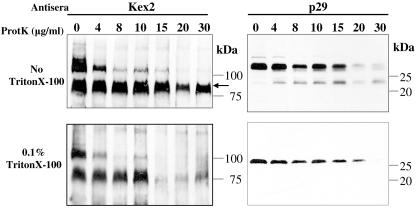
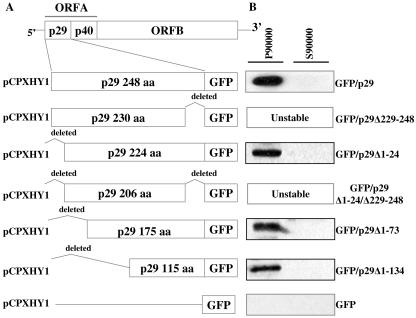
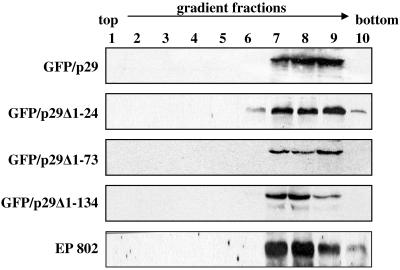
The mycovirus cryphonectria hypovirus 1 (CHV1) causes proliferation of vesicles in its host, Cryphonectria parasitica, the causal agent of chestnut blight. These vesicles have previously been shown to contain both CHV1 genomic double-stranded RNA (dsRNA) and RNA polymerase activity. To determine the cellular origins of these virus-induced membrane structures, we compared the fractionation of several cellular and viral markers. Results showed that viral dsRNA, helicase, polymerase, and protease p29 copurify with C. parasitica trans-Golgi network (TGN) markers, suggesting that the virus utilizes the fungal TGN for replication. We also show that the CHV1 protease p29 associates with vesicle membranes and is resistant to treatments that would release peripheral membrane proteins. Thus, p29 behaves as an integral membrane protein of the vesicular fraction derived from the fungal TGN. Protease p29 was also found to be fully susceptible to proteolytic digestion in the absence of detergent and, thus, is wholly or predominantly on the cytoplasmic face of the vesicles. Fractionation analysis of p29 deletion variants showed that sequences in the C terminal of p29 mediate membrane association. In particular, the C-terminal portion of the protein (Met-135-Gly-248) is sufficient for membrane association and is enough to direct p29 to the TGN vesicles in the absence of other viral elements.
Figures
Similar articles
-
Hypovirus papain-like protease p29 functions in trans to enhance viral double-stranded RNA accumulation and vertical transmission.J Virol. 2003 Nov;77(21):11697-707. doi: 10.1128/jvi.77.21.11697-11707.2003. J Virol. 2003. PMID: 14557655 Free PMC article.
-
The mycovirus CHV1 disrupts secretion of a developmentally regulated protein in Cryphonectria parasitica.J Virol. 2012 Jun;86(11):6067-74. doi: 10.1128/JVI.05756-11. Epub 2012 Mar 21. J Virol. 2012. PMID: 22438560 Free PMC article.
-
Synergism between a mycoreovirus and a hypovirus mediated by the papain-like protease p29 of the prototypic hypovirus CHV1-EP713.J Gen Virol. 2006 Dec;87(Pt 12):3703-3714. doi: 10.1099/vir.0.82213-0. J Gen Virol. 2006. PMID: 17098988
-
Cryphonectria parasitica, the causal agent of chestnut blight: invasion history, population biology and disease control.Mol Plant Pathol. 2018 Jan;19(1):7-20. doi: 10.1111/mpp.12542. Epub 2017 Apr 24. Mol Plant Pathol. 2018. PMID: 28142223 Free PMC article. Review.
-
[Cryphonectria parasitica as a host of fungal viruses: a tool useful to unravel the mycovirus world].Uirusu. 2014;64(1):11-24. doi: 10.2222/jsv.64.11. Uirusu. 2014. PMID: 25765976 Review. Japanese.
Cited by
-
Hypovirus infection induces proliferation and perturbs functions of mitochondria in the chestnut blight fungus.Front Microbiol. 2023 Jun 28;14:1206603. doi: 10.3389/fmicb.2023.1206603. eCollection 2023. Front Microbiol. 2023. PMID: 37448575 Free PMC article.
-
Symptomatic plant viroid infections in phytopathogenic fungi.Proc Natl Acad Sci U S A. 2019 Jun 25;116(26):13042-13050. doi: 10.1073/pnas.1900762116. Epub 2019 Jun 10. Proc Natl Acad Sci U S A. 2019. PMID: 31182602 Free PMC article.
-
Mycoviruses of an endophytic fungus can replicate in plant cells: evolutionary implications.Sci Rep. 2017 May 15;7(1):1908. doi: 10.1038/s41598-017-02017-3. Sci Rep. 2017. PMID: 28507331 Free PMC article.
-
New Detection Methods for Cryphonectria Hypovirus 1 (CHV1) through SYBR Green-Based Real-Time PCR and Loop-Mediated Isothermal Amplification (LAMP).Viruses. 2024 Jul 26;16(8):1203. doi: 10.3390/v16081203. Viruses. 2024. PMID: 39205177 Free PMC article.
-
CpATG8, a Homolog of Yeast Autophagy Protein ATG8, Is Required for Pathogenesis and Hypovirus Accumulation in the Chest Blight Fungus.Front Cell Infect Microbiol. 2019 Jul 10;9:222. doi: 10.3389/fcimb.2019.00222. eCollection 2019. Front Cell Infect Microbiol. 2019. PMID: 31355148 Free PMC article.
References
-
- Bienz, K., D. Egger, and L. Pasamontes. 1987. Association of polioviral proteins of the P2 genomic region with the viral replication complex and virus-induced membrane synthesis as visualized by electron microscopic immunocytochemistry and autoradiography. Virology 160:220-226. - PubMed
-
- Bordier, C. 1981. Phase separation of integral membrane proteins in Triton X-114. J. Biol. Chem. 256:1604-1607. - PubMed
-
- Chen, B., G. H. Choi, and D. L. Nuss. 1994. Attenuation of fungal virulence by synthetic infectious hypovirus transcripts. Science 264:1762-1764. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Miscellaneous
