

Identification of specific calcitonin-like receptor residues important for calcitonin gene-related peptide high affinity binding
- PMID: 16776831
- PMCID: PMC1525162
- DOI: 10.1186/1471-2210-6-9
Identification of specific calcitonin-like receptor residues important for calcitonin gene-related peptide high affinity binding
Abstract
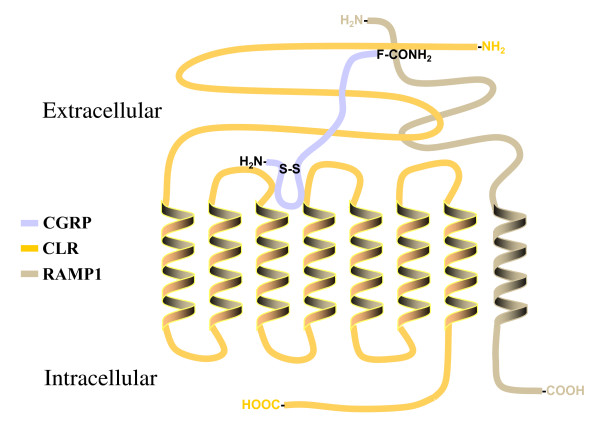
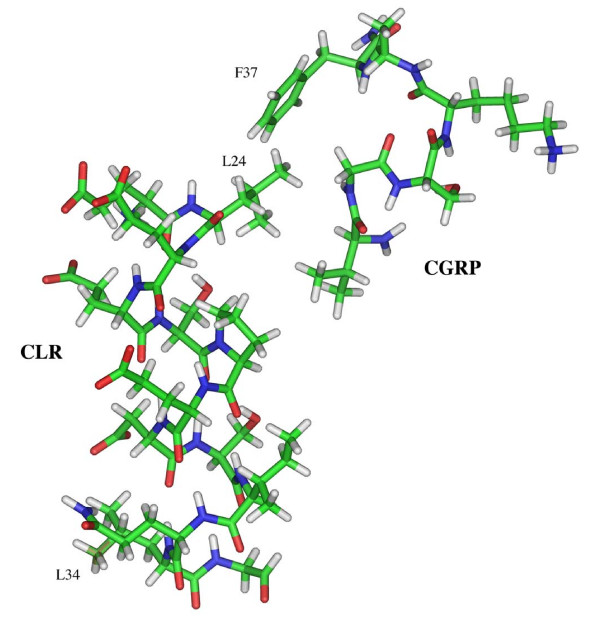
Background: Calcitonin gene-related peptide (CGRP) is a vasoactive neuropeptide whose biological activity has potential therapeutic value for many vascular related diseases. CGRP is a 37 amino acid neuropeptide that signals through a G protein-coupled receptor belonging to the secretin receptor family. Previous studies on the calcitonin-like receptor (CLR), which requires co-expression of the receptor-activity-modifying protein-1 (RAMP1) to function as a CGRP receptor, have shown an 18 amino acid N-terminus sequence important for binding CGRP. Moreover, several investigations have recognized the C-terminal amidated phenylalanine (F37) of CGRP as essential for docking to the mature receptor. Therefore, we hypothesize that hydrophobic amino acids within the previously characterized 18 amino acid CLR N-terminus domain are important binding contacts for the C-terminal phenylalaninamide of CGRP.
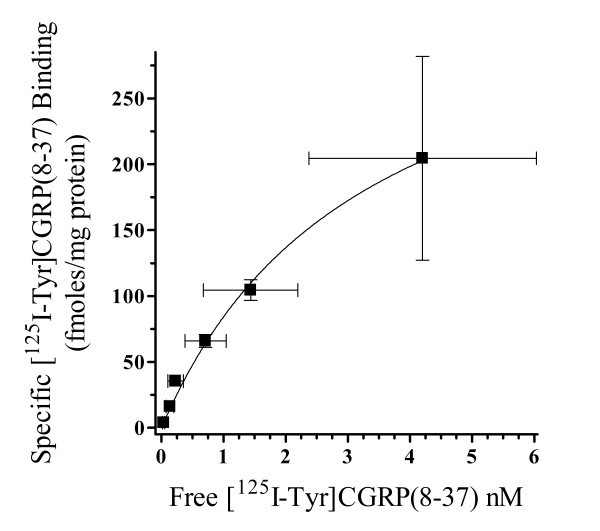
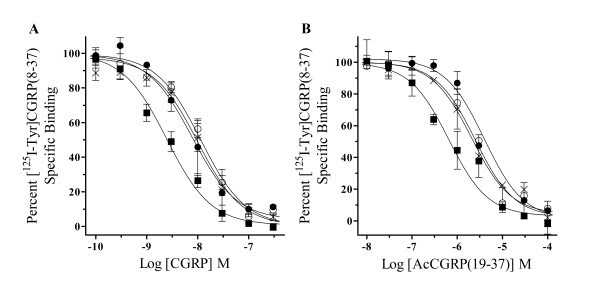
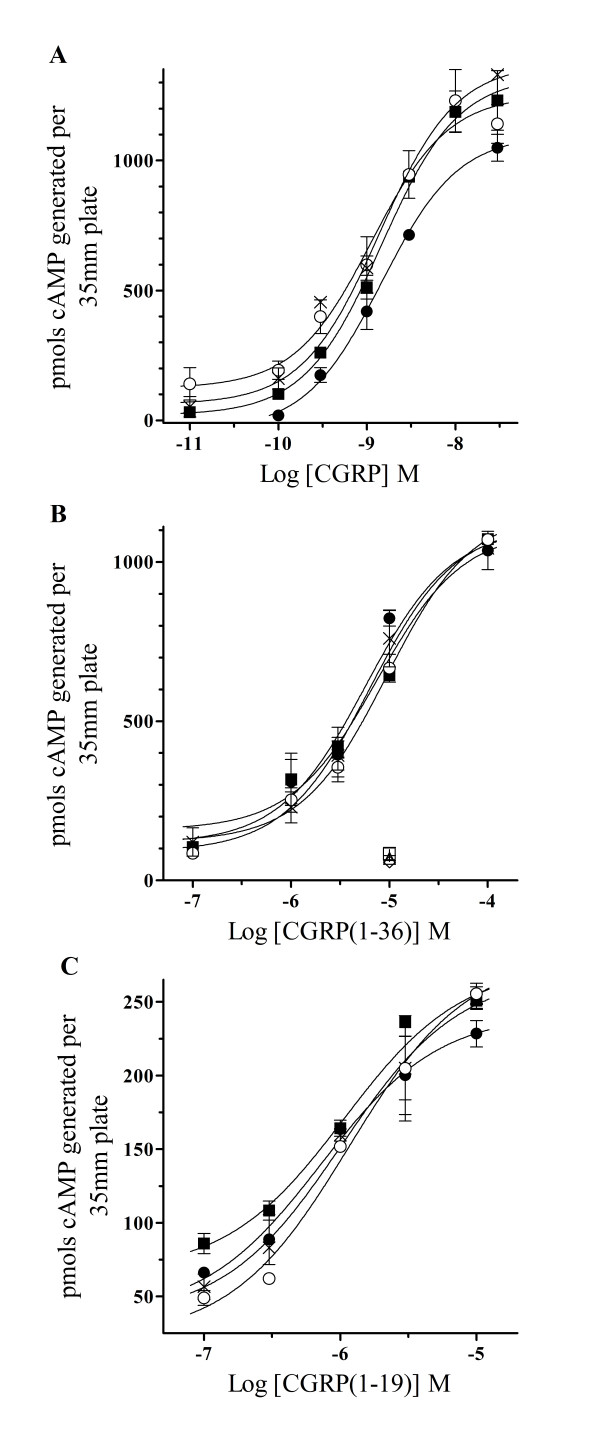
Results: Two leucine residues within this previously characterized CLR N-terminus domain, when mutated to alanine and expressed on HEK293T cells stably transfected with RAMP1, demonstrated a significantly decreased binding affinity for CGRP compared to wild type receptor. Additional decreases in binding affinity for CGRP were not found when both leucine mutations were expressed in the same CLR construct. Decreased binding characteristic of these leucine mutant receptors was observed for all CGRP ligands tested that contained the necessary amidated phenylalanine at their C-terminus. However, there was no difference in the potency of CGRP to increase cAMP production by these leucine mutant receptors when compared to wild type CLR, consistent with the notion that the neuropeptide C-terminal F37 is important for docking but not activation of the receptor. This observation was conserved when modified CGRP ligands lacking the amidated F37 demonstrated similar potencies to generate cAMP at both wild type and mutant CLRs. Furthermore, these modified CGRP ligands displayed a significant but similar loss of binding for all leucine mutant and wild type CLR because the important receptor contact on the neuropeptide was missing in all experimental situations.
Conclusion: These results are consistent with previous structure-function investigations of the neuropeptide and are the first to propose specific CLR binding contacts for the amidated F37 of CGRP that are important for docking but not activation of the mature CGRP receptor.
Figures
References
-
- Gulbenkian S, Saetrum OO, Ekman R, Costa AN, Wharton J, Polak JM, Melo Queiroz e, Edvinsson L. Peptidergic innervation of human epicardial coronary arteries. Circ Res. 1993;73:579–588. - PubMed
-
- Edvinsson L, Sams A, Jansen-Olesen I, Tajti J, Kane SA, Rutledge RZ, Koblan KS, Hill RG, Longmore J. Characterisation of the effects of a non-peptide CGRP receptor antagonist in SK-N-MC cells and isolated human cerebral arteries. Eur J Pharmacol. 2001;415:39–44. doi: 10.1016/S0014-2999(00)00934-1. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
