

Sarcoplasmic reticulum Ca2+ release and depletion fail to affect sarcolemmal ion channel activity in mouse skeletal muscle
- PMID: 16777939
- PMCID: PMC1819412
- DOI: 10.1113/jphysiol.2006.112367
Sarcoplasmic reticulum Ca2+ release and depletion fail to affect sarcolemmal ion channel activity in mouse skeletal muscle
Abstract
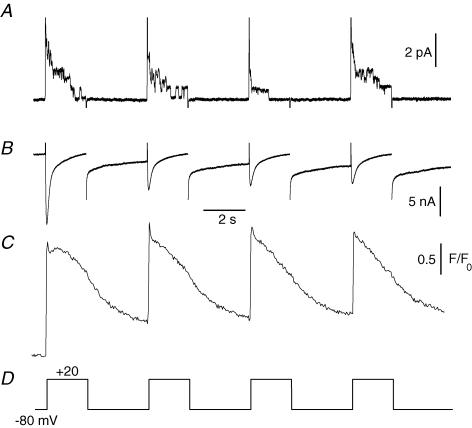
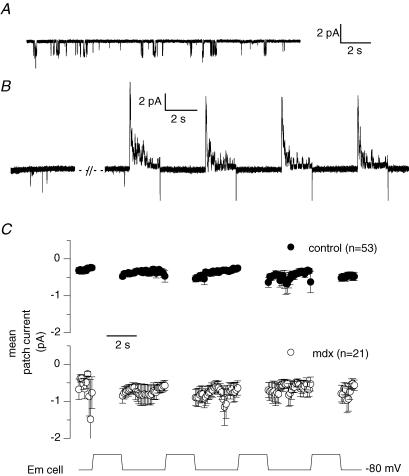
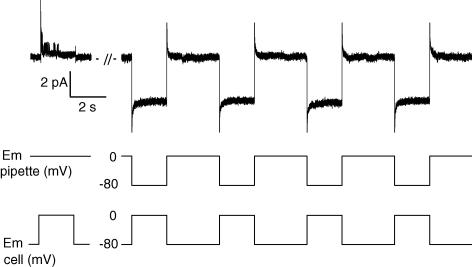
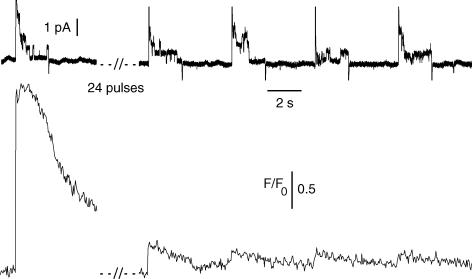
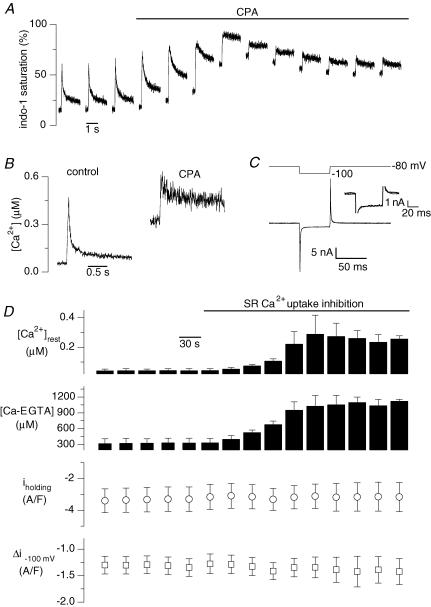
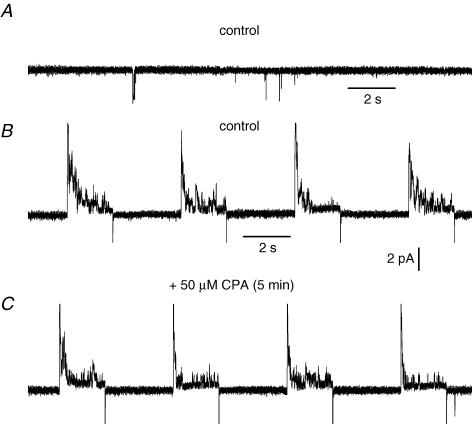
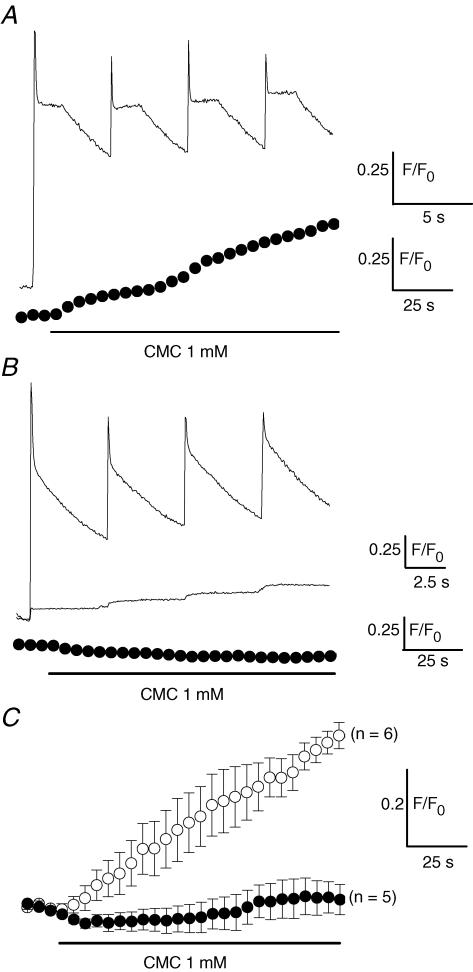
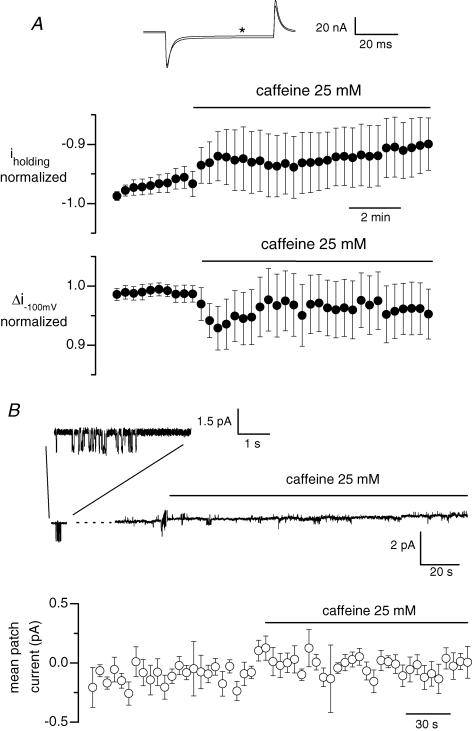
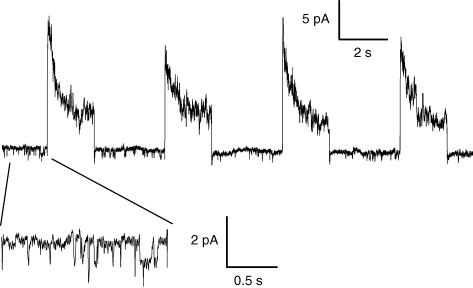
In skeletal muscle, sarcoplasmic reticulum (SR) Ca2+ depletion is suspected to trigger a calcium entry across the plasma membrane and recent studies also suggest that the opening of channels spontaneously active at rest and possibly involved in Duchenne dystrophy may be regulated by SR Ca2+ depletion. Here we simultaneously used the cell-attached and whole-cell voltage-clamp techniques as well as intracellular Ca2+ measurements on single isolated mouse skeletal muscle fibres to unravel any possible change in membrane conductance that would depend upon SR Ca2+ release and/or SR Ca2+ depletion. Delayed rectifier K+ single channel activity was routinely detected during whole-cell depolarizing pulses. In addition the activity of channels carrying unitary inward currents of approximately 1.5 pA at -80 mV was detected in 17 out of 127 and in 21 out of 59 patches in control and mdx dystrophic fibres, respectively. In both populations of fibres, large whole-cell depolarizing pulses did not reproducibly increase this channel activity. This was also true when, repeated application of the whole-cell pulses led to exhaustion of the Ca2+ transient. SR Ca2+ depletion produced by the SR Ca2+ pump inhibitor cyclopiazonic acid (CPA) also failed to induce any increase in the resting whole-cell conductance and in the inward single channel activity. Overall results indicate that voltage-activated SR Ca2+ release and/or SR Ca2+ depletion are not sufficient to activate the opening of channels carrying inward currents at negative voltages and challenge the physiological relevance of a store-operated membrane conductance in adult skeletal muscle.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous