

MBD2 is a critical component of a methyl cytosine-binding protein complex isolated from primary erythroid cells
- PMID: 16778143
- PMCID: PMC1895583
- DOI: 10.1182/blood-2006-04-016394
MBD2 is a critical component of a methyl cytosine-binding protein complex isolated from primary erythroid cells
Abstract
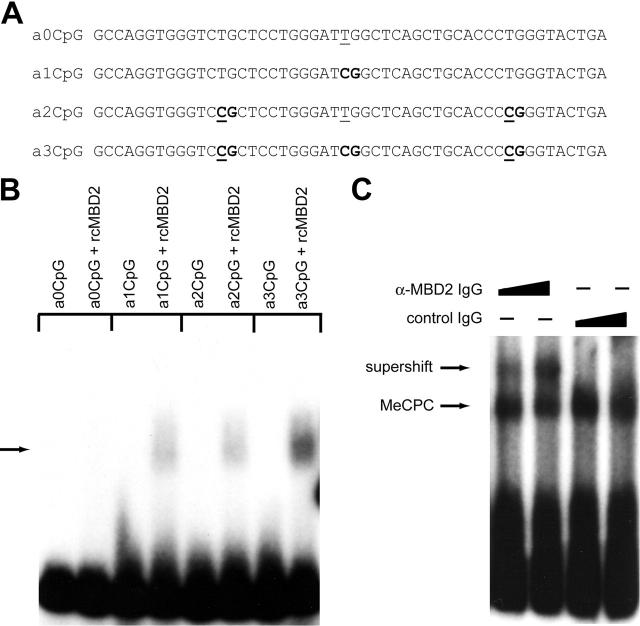
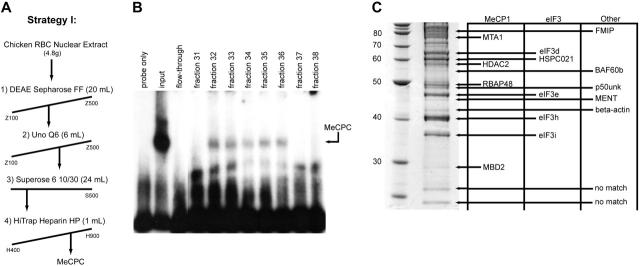
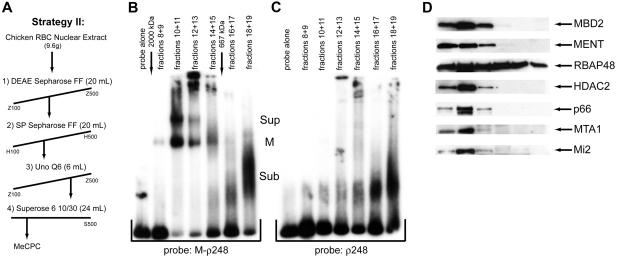
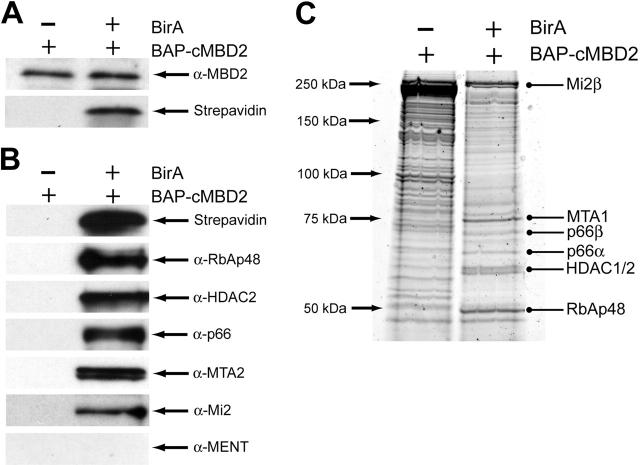
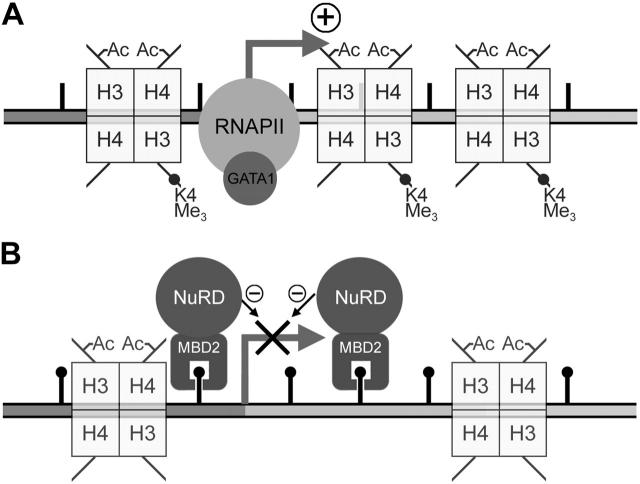
The chicken embryonic beta-type globin gene, rho, is a member of a small group of vertebrate genes whose developmentally regulated expression is mediated by DNA methylation. Previously, we have shown that a methyl cytosine-binding complex binds to the methylated rho-globin gene in vitro. We have now chromatographically purified and characterized this complex from adult chicken primary erythroid cells. Four components of the MeCP1 transcriptional repression complex were identified: MBD2, RBAP48, HDAC2, and MTA1. These 4 proteins, as well as the zinc-finger protein p66 and the chromatin remodeling factor Mi2, were found to coelute by gel-filtration analysis and pull-down assays. We conclude that these 6 proteins are components of the MeCPC. In adult erythrocytes, significant enrichment for MBD2 is seen at the inactive rho-globin gene by chromatin immunoprecipitation assay, whereas no enrichment is observed at the active beta(A)-globin gene, demonstrating MBD2 binds to the methylated and transcriptionally silent rho-globin gene in vivo. Knock-down of MBD2 resulted in up-regulation of a methylated rho-gene construct in mouse erythroleukemic (MEL)-rho cells. These results represent the first purification of a MeCP1-like complex from a primary cell source and provide support for a role for MBD2 in developmental gene regulation.
Figures
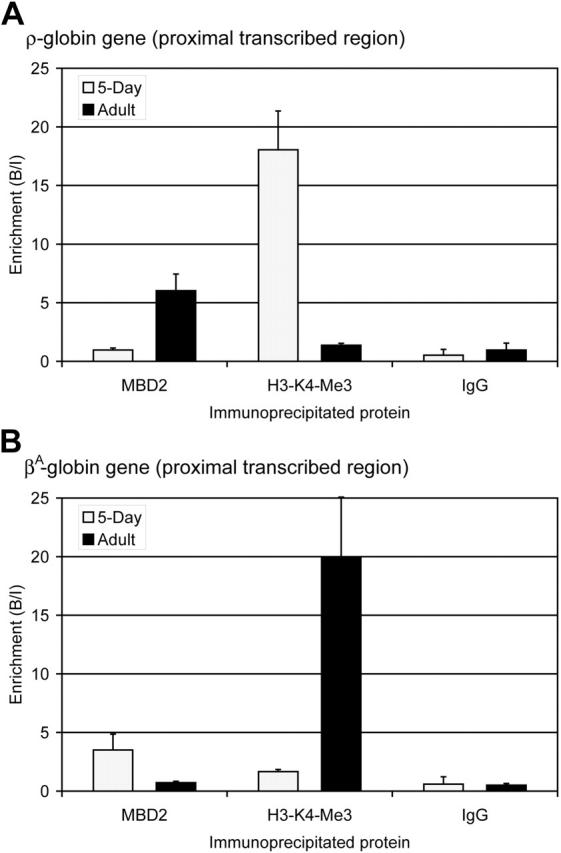
 ) and adult (▪) erythrocytes as determined by ChIP assay. The data show that MBD2 is depleted from the transcriptionally active ρ-globin gene in 5-day erythrocytes but enriched at the transcriptionally inactive and methylated ρ-globin gene in adult erythrocytes. In contrast, H3-K4-Me3 is enriched at the transcriptionally active ρ-globin gene and depleted at the transcriptionally inactive gene. No enrichment was seen using anti-rabbit IgG in either 5-day or adult erythrocytes, verifying the interaction of these specific proteins with the ρ-globin gene. The data represent the average of 3 independent experiments, with the SD indicated by the bar. (B) Enrichment for MBD2, H3-K4-Me3, and IgG at the βA-globin gene in 5-day () and adult (▪) erythrocytes as determined by ChIP assay. The data show that MBD2 is enriched at the transcriptionally inactive and methylated βA-globin gene in 5-day erythrocytes but depleted from the transcriptionally active βA-globin gene in adult erythrocytes. Once again, H3-K4-Me3 is depleted from the transcriptionally inactive βA-globin gene and enriched at the transcriptionally active gene. No enrichment was seen using anti-rabbit IgG in either 5-day or adult erythrocytes, verifying the interaction of these specific proteins with the βA-globin gene. The data represent the average of 3 independent experiments, with the SD indicated by the bar.
) and adult (▪) erythrocytes as determined by ChIP assay. The data show that MBD2 is depleted from the transcriptionally active ρ-globin gene in 5-day erythrocytes but enriched at the transcriptionally inactive and methylated ρ-globin gene in adult erythrocytes. In contrast, H3-K4-Me3 is enriched at the transcriptionally active ρ-globin gene and depleted at the transcriptionally inactive gene. No enrichment was seen using anti-rabbit IgG in either 5-day or adult erythrocytes, verifying the interaction of these specific proteins with the ρ-globin gene. The data represent the average of 3 independent experiments, with the SD indicated by the bar. (B) Enrichment for MBD2, H3-K4-Me3, and IgG at the βA-globin gene in 5-day () and adult (▪) erythrocytes as determined by ChIP assay. The data show that MBD2 is enriched at the transcriptionally inactive and methylated βA-globin gene in 5-day erythrocytes but depleted from the transcriptionally active βA-globin gene in adult erythrocytes. Once again, H3-K4-Me3 is depleted from the transcriptionally inactive βA-globin gene and enriched at the transcriptionally active gene. No enrichment was seen using anti-rabbit IgG in either 5-day or adult erythrocytes, verifying the interaction of these specific proteins with the βA-globin gene. The data represent the average of 3 independent experiments, with the SD indicated by the bar.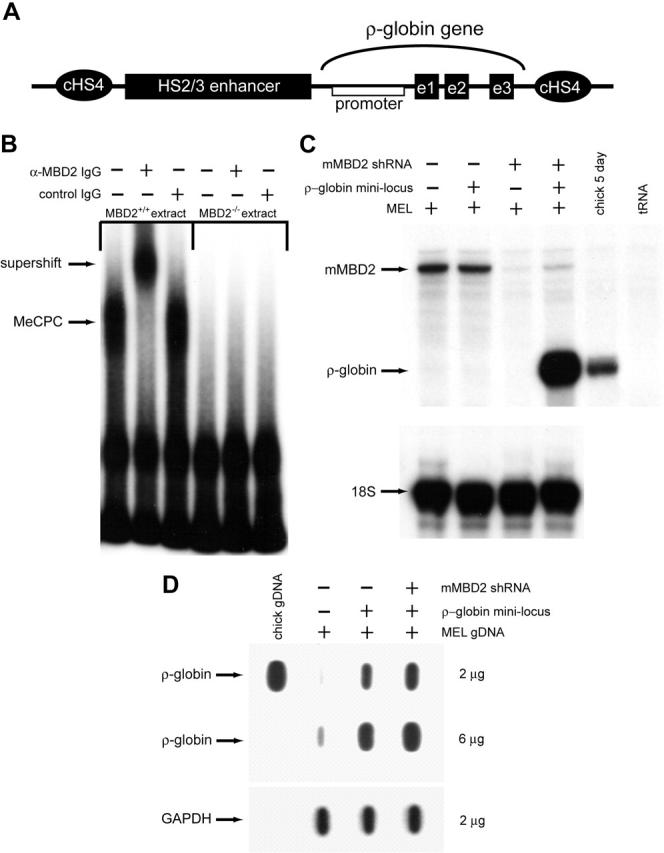
References
-
- McGhee JD, Ginder GD. Specific DNA methylation sites in the vicinity of the chicken beta-globin genes. Nature. 1979;280: 419-420. - PubMed
-
- van der Ploeg LH, Flavell RA. DNA methylation in the human gamma delta beta-globin locus in erythroid and nonerythroid tissues. Cell. 1980;19: 947-958. - PubMed
-
- Groudine M, Weintraub H. Activation of globin genes during chicken development. Cell. 1981; 24: 393-401. - PubMed
-
- Herman JG, Baylin SB. Gene silencing in cancer in association with promoter hypermethylation. N Engl J Med. 2003;349: 2042-2054. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
