

Toxoplasma gondii scavenges host-derived lipoic acid despite its de novo synthesis in the apicoplast
- PMID: 16778769
- PMCID: PMC1500979
- DOI: 10.1038/sj.emboj.7601189
Toxoplasma gondii scavenges host-derived lipoic acid despite its de novo synthesis in the apicoplast
Abstract
In contrast to other eukaryotes, which manufacture lipoic acid, an essential cofactor for several vital dehydrogenase complexes, within the mitochondrion, we show that the plastid (apicoplast) of the obligate intracellular protozoan parasite Toxoplasma gondii is the only site of de novo lipoate synthesis. However, antibodies specific for protein-attached lipoate reveal the presence of lipoylated proteins in both, the apicoplast and the mitochondrion of T. gondii. Cultivation of T. gondii-infected cells in lipoate-deficient medium results in substantially reduced lipoylation of mitochondrial (but not apicoplast) proteins. Addition of exogenous lipoate to the medium can rescue this effect, showing that the parasite scavenges this cofactor from the host. Exposure of T. gondii to lipoate analogues in lipoate-deficient medium leads to growth inhibition, suggesting that T. gondii might be auxotrophic for this cofactor. Phylogenetic analyses reveal the secondary loss of the mitochondrial lipoate synthase gene after the acquisition of the plastid. Our studies thus reveal an unexpected metabolic deficiency in T. gondii and raise the question whether the close interaction of host mitochondria with the parasitophorous vacuole is connected to lipoate supply by the host.
Figures
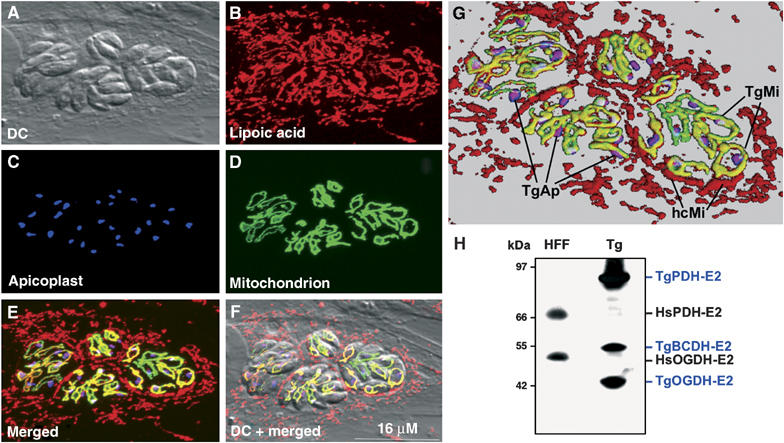

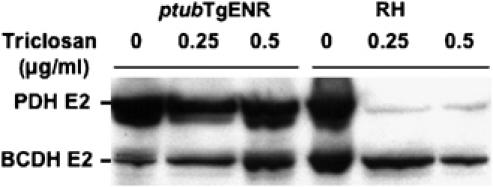
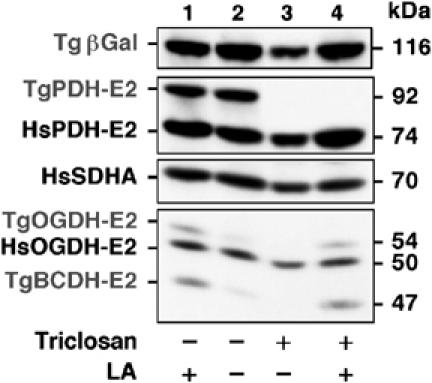
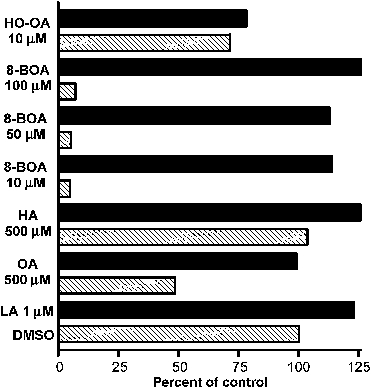

References
-
- Ali ST, Moir AJ, Ashton PR, Engel PC, Guest JR (1990) Octanoylation of the lipoyl domains of the pyruvate dehydrogenase complex in a lipoyl-deficient strain of Escherichia coli. Mol Microbiol 4: 943–950 - PubMed
-
- Andersson SG, Zomorodipour A, Andersson JO, Sicheritz-Ponten T, Alsmark UC, Podowski RM, Naslund AK, Eriksson AS, Winkler HH, Kurland CG (1998) The genome sequence of Rickettsia prowazekii and the origin of mitochondria. Nature 396: 133–140 - PubMed
-
- Augustin S, Nolden M, Müller S, Hardt O, Arnold I, Langer T (2005) Characterization of peptides released from mitochondria: evidence for constant proteolysis and peptide efflux. J Biol Chem 280: 2691–2699 - PubMed
-
- Biewenga G, Haenen G, Bast A (1997) An overview of lipoate chemistry. In Lipoic Acid in Health and Disease, Fuchs J, Packer L, Zimmer G (eds) Vol. 6, pp 1–32. New York: Marcel Dekker
-
- Coppens I, Joiner KA (2001) Parasite–host cell interactions in toxoplasmosis: new avenues for intervention? Expert Rev Mol Med 2001: 1–20 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
