

Regulation of mitochondrial morphology through proteolytic cleavage of OPA1
- PMID: 16778770
- PMCID: PMC1500981
- DOI: 10.1038/sj.emboj.7601184
Regulation of mitochondrial morphology through proteolytic cleavage of OPA1
Abstract
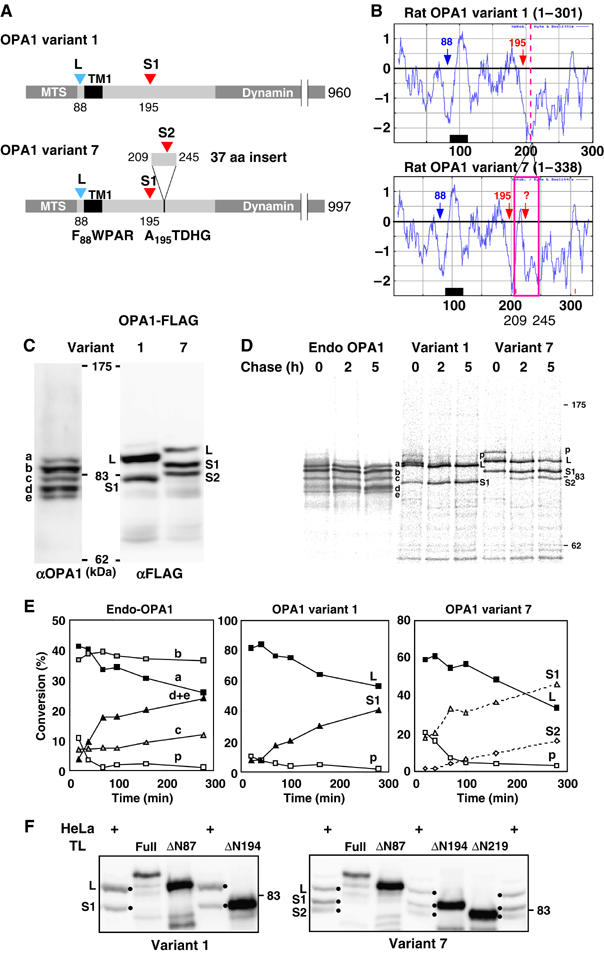
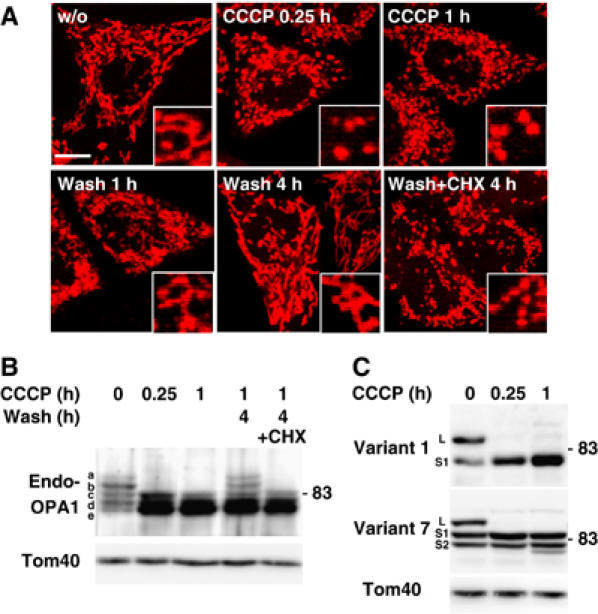
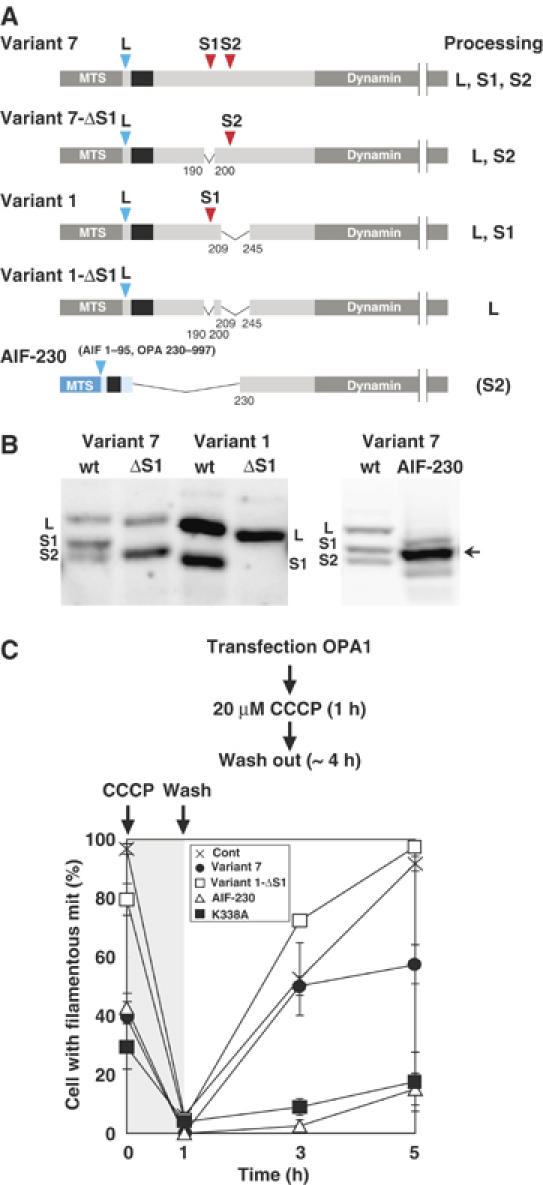
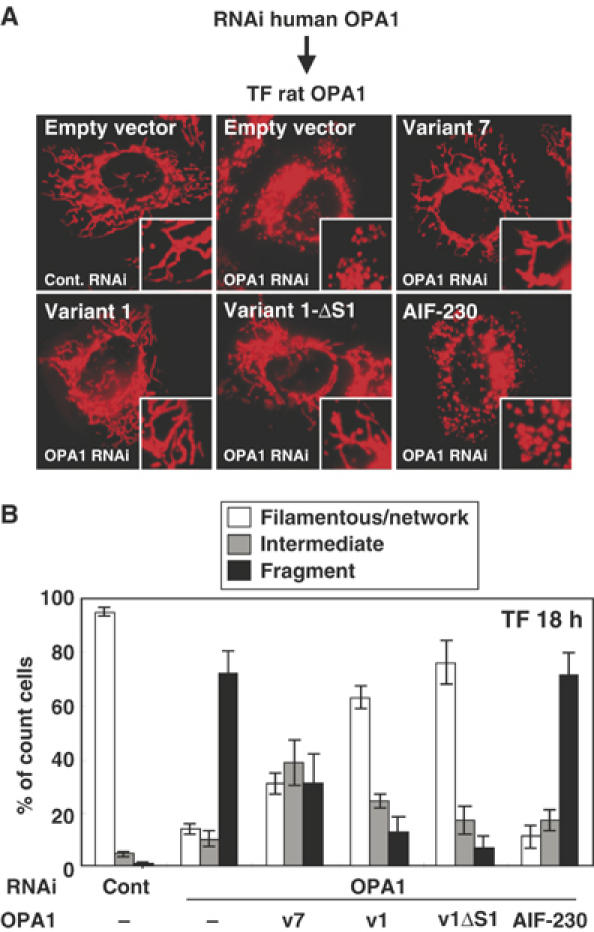
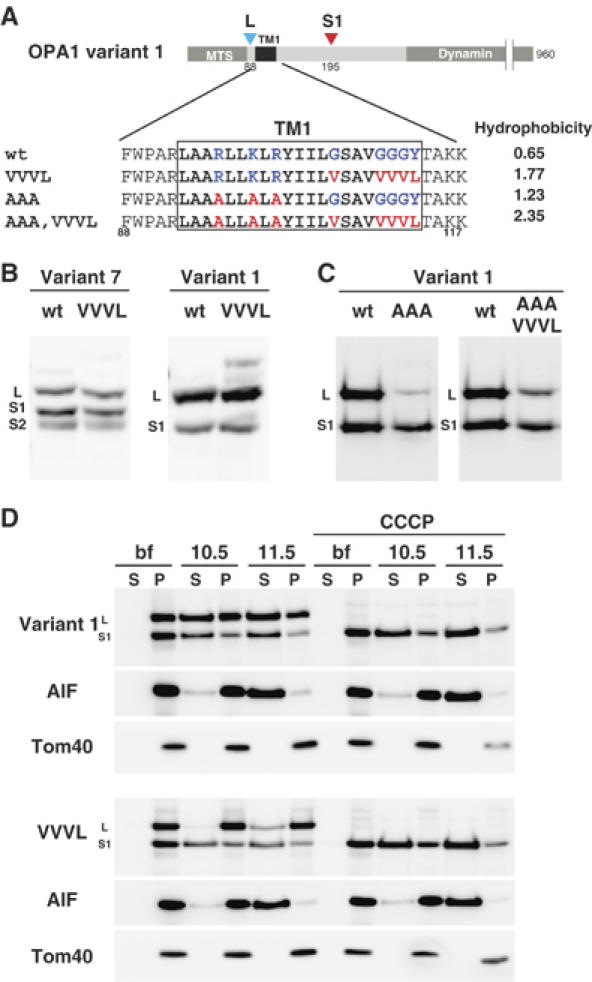
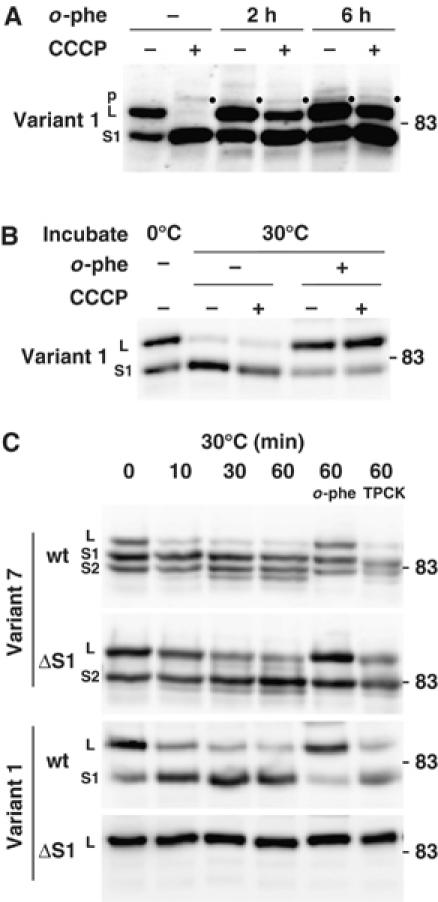
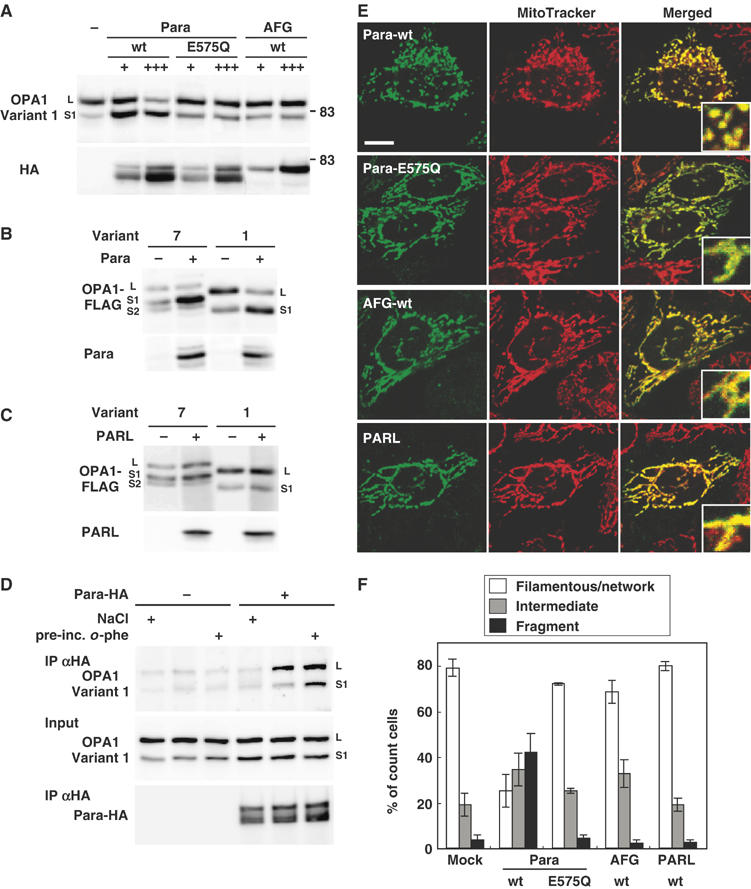
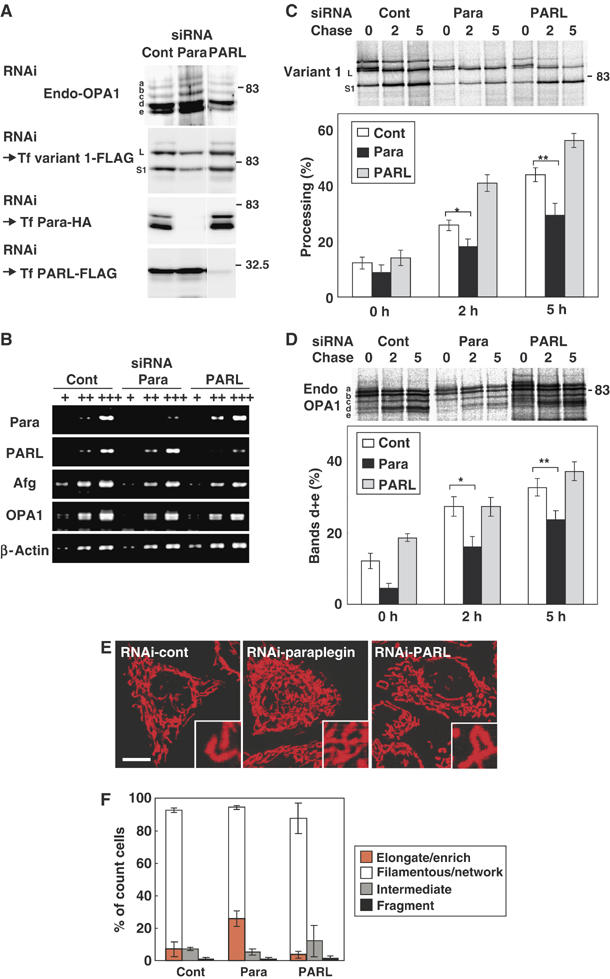
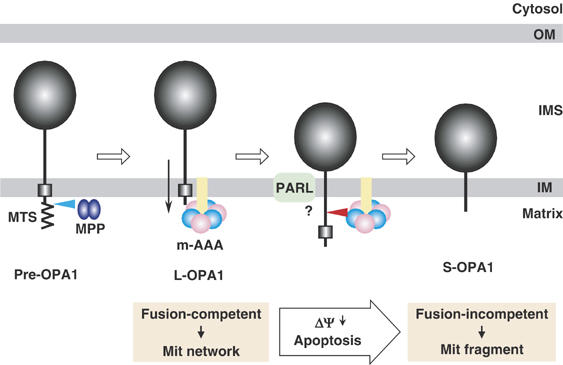
The dynamin-like GTPase OPA1, a causal gene product of human dominant optic atrophy, functions in mitochondrial fusion and inner membrane remodeling. It has several splice variants and even a single variant is found as several processed forms, although their functional significance is unknown. In yeast, mitochondrial rhomboid protease regulates mitochondrial function and morphology through proteolytic cleavage of Mgm1, the yeast homolog of OPA1. We demonstrate that OPA1 variants are synthesized with a bipartite-type mitochondrial targeting sequence. During import, the matrix-targeting signal is removed and processed forms (L-isoforms) are anchored to the inner membrane in type I topology. L-isoforms undergo further processing in the matrix to produce S-isoforms. Knockdown of OPA1 induced mitochondrial fragmentation, whose network morphology was recovered by expression of L-isoform but not S-isoform, indicating that only L-isoform is fusion-competent. Dissipation of membrane potential, expression of m-AAA protease paraplegin, or induction of apoptosis stimulated this processing along with the mitochondrial fragmentation. Thus, mammalian mitochondrial function and morphology is regulated through processing of OPA1 in a DeltaPsi-dependent manner.
Figures
References
-
- Alexander C, Votruba M, Pesch UE, Thiselton DL, Mayer S, Moore A, Rodriguez M, Kellner U, Leo-Kottler B, Auburger G, Bhattacharya SS, Wissinger B (2000) OPA1, encoding a dynamin-related GTPase, is mutated in autosomal dominant optic atrophy linked to chromosome 3q28. Nat Genet 26: 211–215 - PubMed
-
- Arlt H, Tauer R, Feldmann H, Neupert W, Langer T (1996) The YTA10-12 complex, an AAA protease with chaperone-like activity in the inner membrane of mitochondria. Cell 85: 875–885 - PubMed
-
- Casari G, De Fusco M, Ciarmatori S, Zeviani M, Mora M, Fernandez P, De Michele G, Filla A, Cocozza S, Marconi R, Durr A, Fontaine B, Ballabio A (1998) Spastic paraplegia and OXPHOS impairment caused by mutations in paraplegin, a nuclear-encoded mitochondrial metalloprotease. Cell 93: 973–983 - PubMed
-
- Chen H, Chan DC (2004) Mitochondrial dynamics in mammals. Curr Top Dev Biol 59: 119–144 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
