

The IL-21 receptor augments Th2 effector function and alternative macrophage activation
- PMID: 16778988
- PMCID: PMC1479424
- DOI: 10.1172/JCI27727
The IL-21 receptor augments Th2 effector function and alternative macrophage activation
Abstract
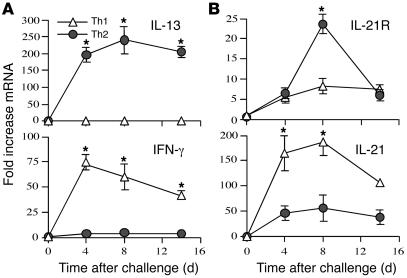
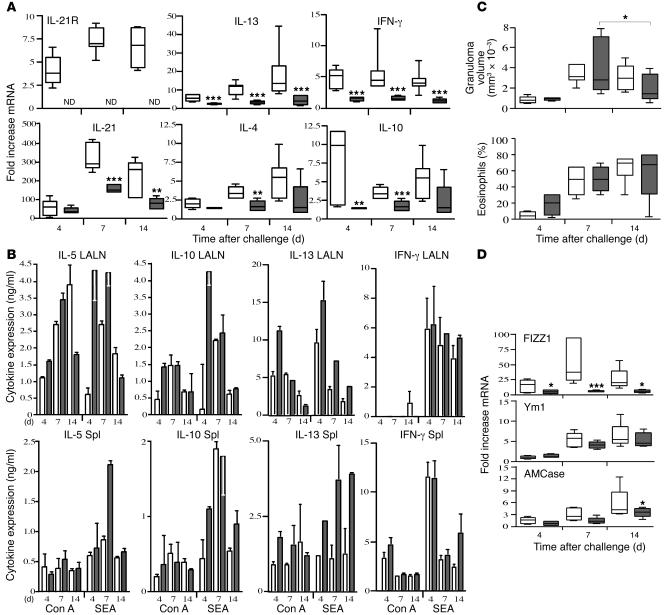
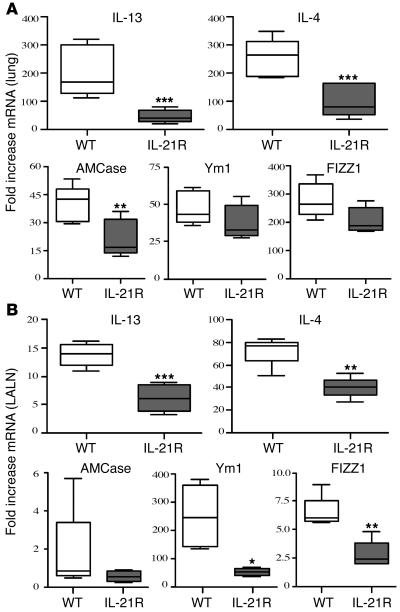
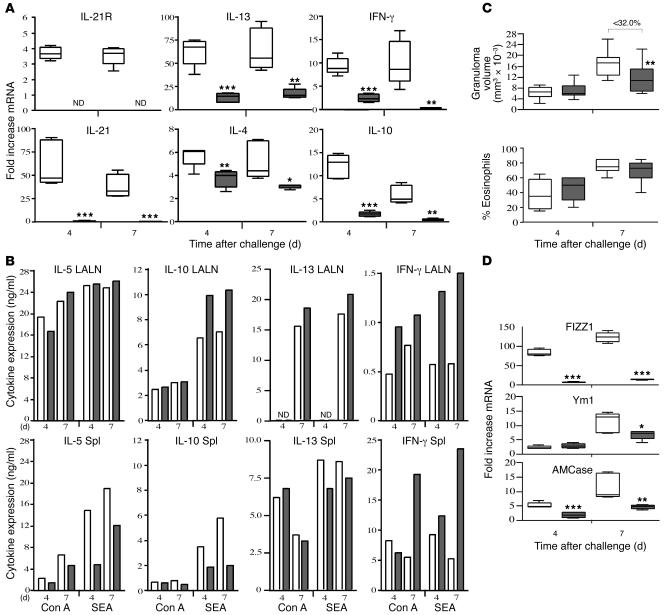
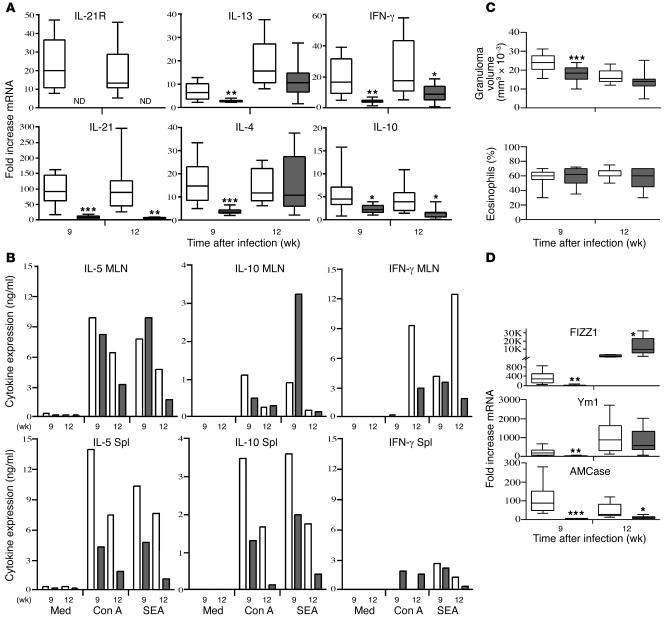
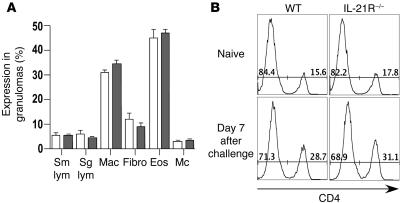
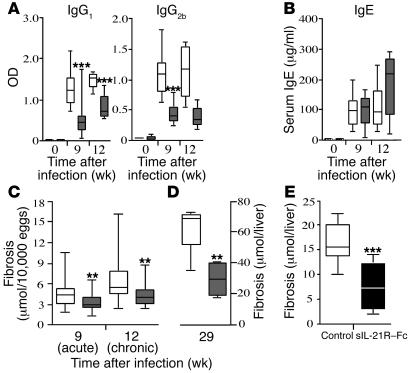
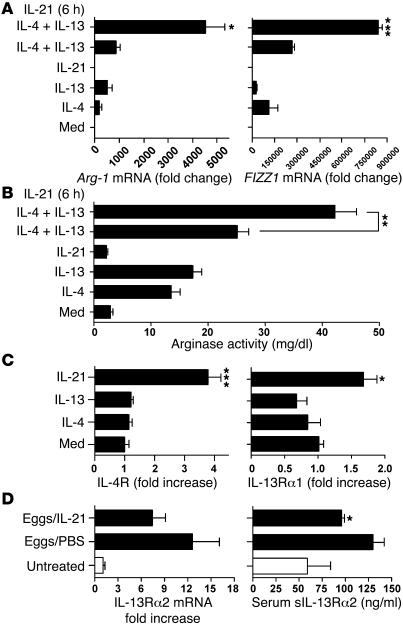
The IL-21 receptor (IL-21R) shows significant homology with the IL-4R, and CD4+ Th2 cells are an important source of IL-21. Here we examined whether the IL-21R regulates the development of Th2 responses in vivo. To do this, we infected IL-21R-/- mice with the Th2-inducing pathogens Schistosoma mansoni and Nippostrongylus brasiliensis and examined the influence of IL-21R deficiency on the development of Th2-dependent pathology. We showed that granulomatous inflammation and liver fibrosis were significantly reduced in S. mansoni-infected IL-21R-/- mice and in IL-21R+/+ mice treated with soluble IL-21R-Fc (sIL-21R-Fc). The impaired granulomatous response was also associated with a marked reduction in Th2 cytokine expression and function, as evidenced by the attenuated IL-4, IL-13, AMCase, Ym1, and FIZZ1 (also referred to as RELMalpha) responses in the tissues. A similarly impaired Th2 response was observed following N. brasiliensis infection. In vitro, IL-21 significantly augmented IL-4Ralpha and IL-13Ralpha1 expression in macrophages, resulting in increased FIZZ1 mRNA and arginase-1 activity following stimulation with IL-4 and IL-13. As such, these data identify the IL-21R as an important amplifier of alternative macrophage activation. Collectively, these results illustrate an essential function for the IL-21R in the development of pathogen-induced Th2 responses, which may have relevance in therapies for both inflammatory and chronic fibrotic diseases.
Figures
References
-
- Parrish-Novak J., et al. Interleukin 21 and its receptor are involved in NK cell expansion and regulation of lymphocyte function. Nature. 2000;408:57–63. - PubMed
-
- Habib T., Nelson A., Kaushansky K. IL-21: a novel IL-2-family lymphokine that modulates B, T, and natural killer cell responses. J. Allergy Clin. Immunol. 2003;112:1033–1045. - PubMed
-
- Vosshenrich C.A., Di Santo J.P. Cytokines: IL-21 joins the gamma(c)-dependent network? Curr. Biol. 2001;11:R175–R177. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
