

Variations in the structure of the prelunate gyrus in Old World monkeys
- PMID: 16779809
- PMCID: PMC2837282
- DOI: 10.1002/ar.a.20350
Variations in the structure of the prelunate gyrus in Old World monkeys
Abstract
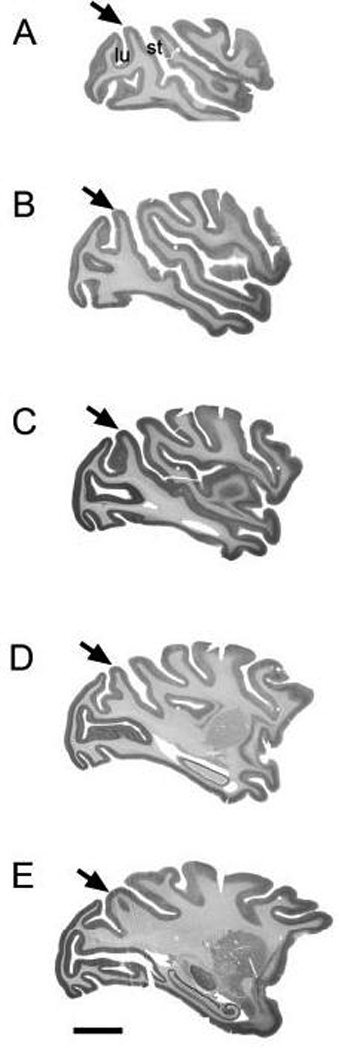
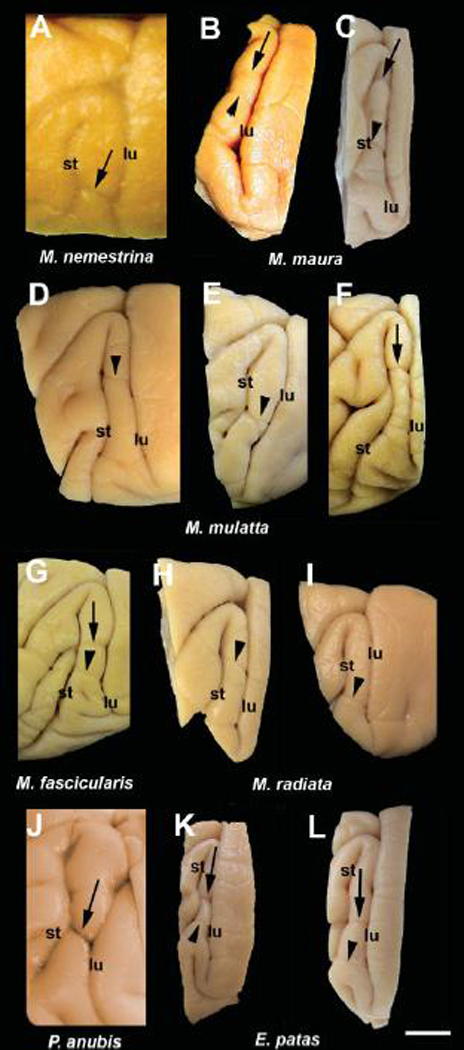
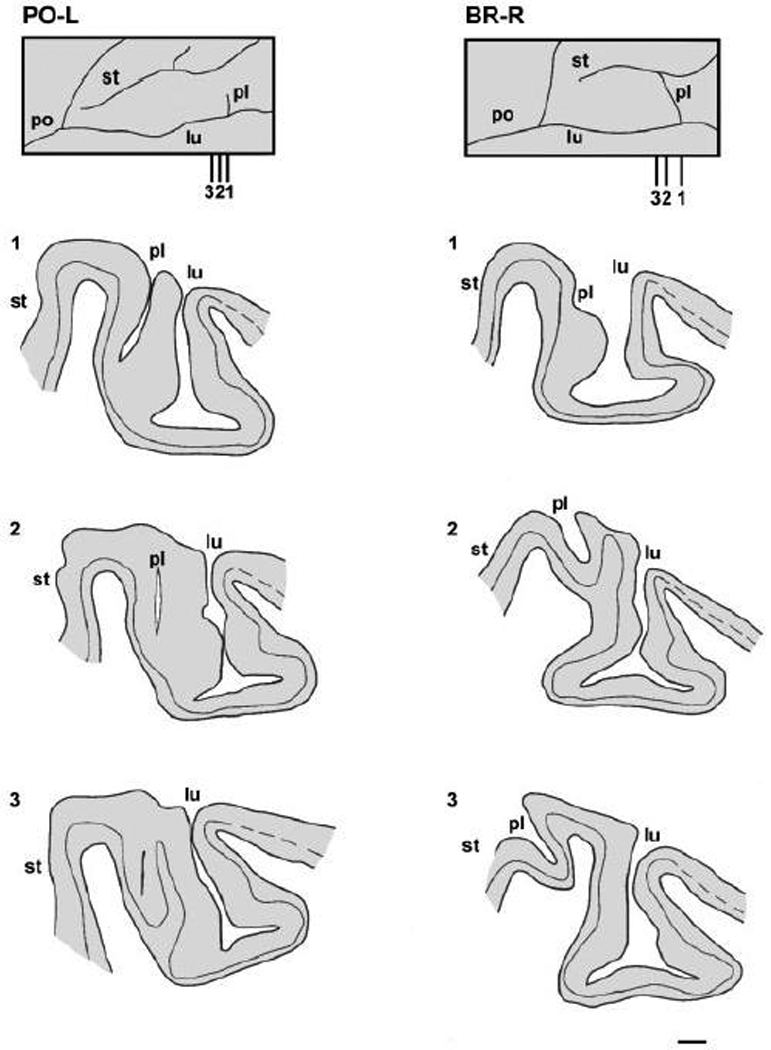
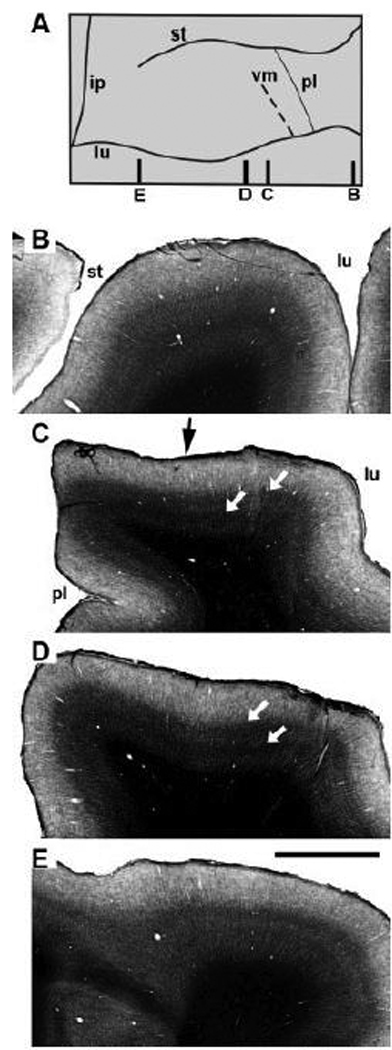
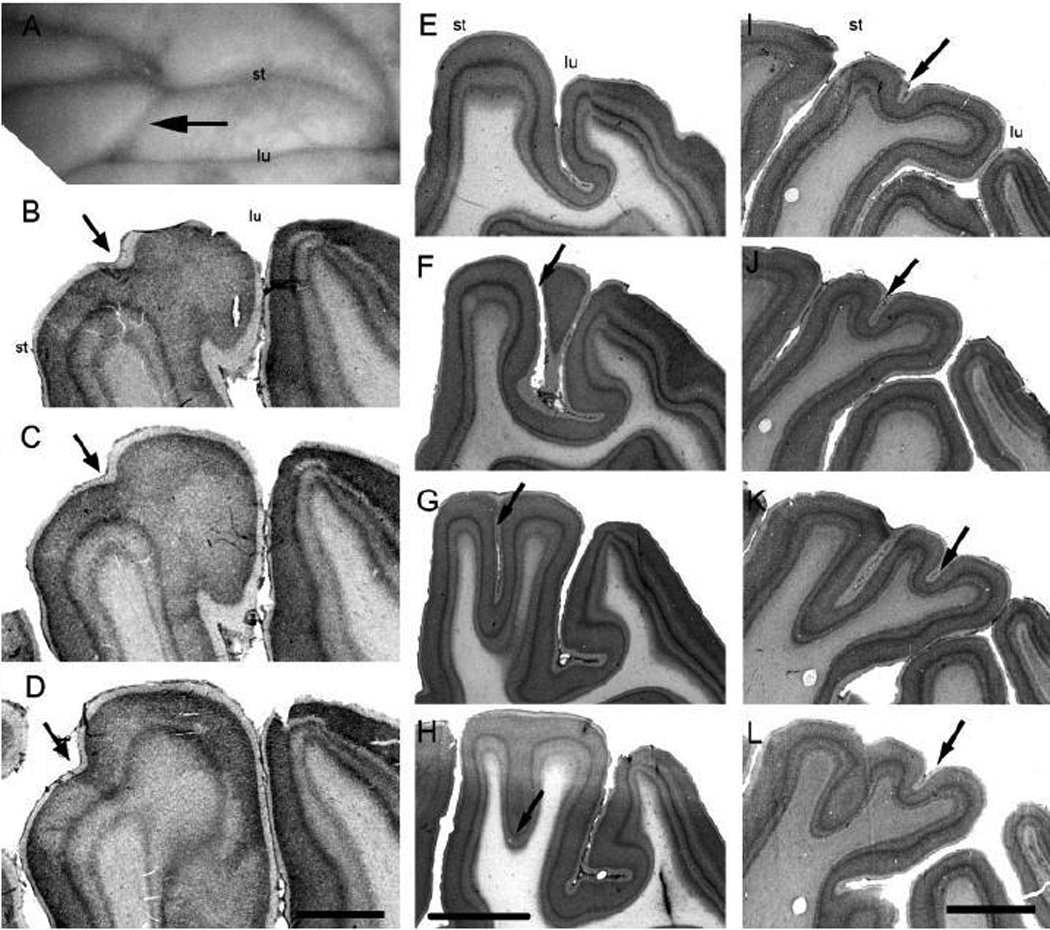
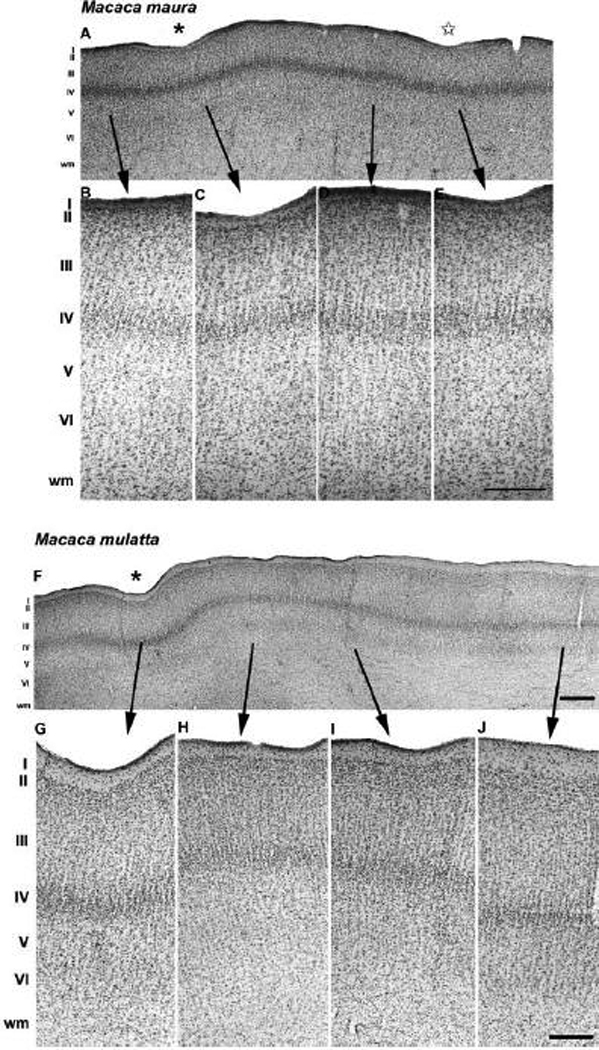
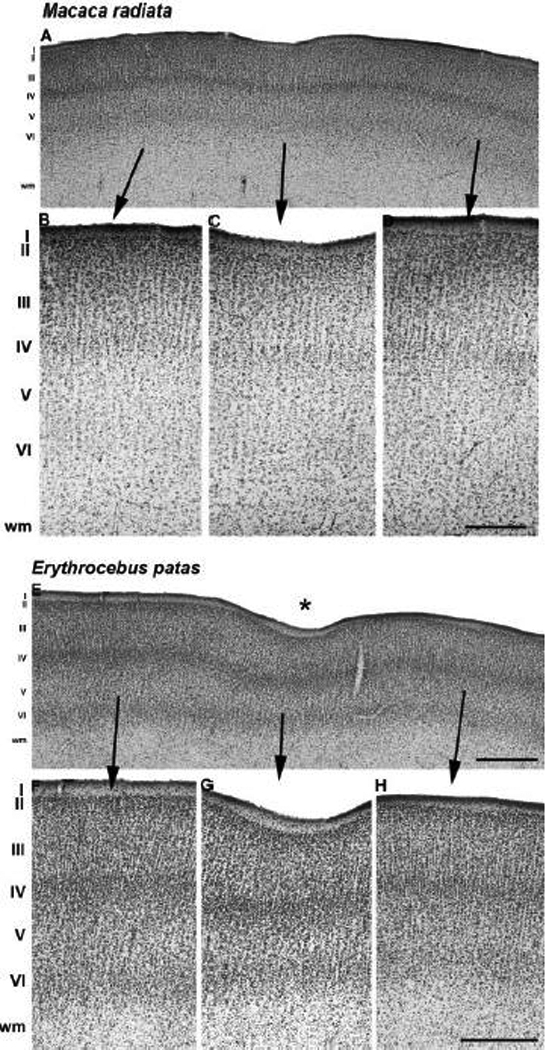
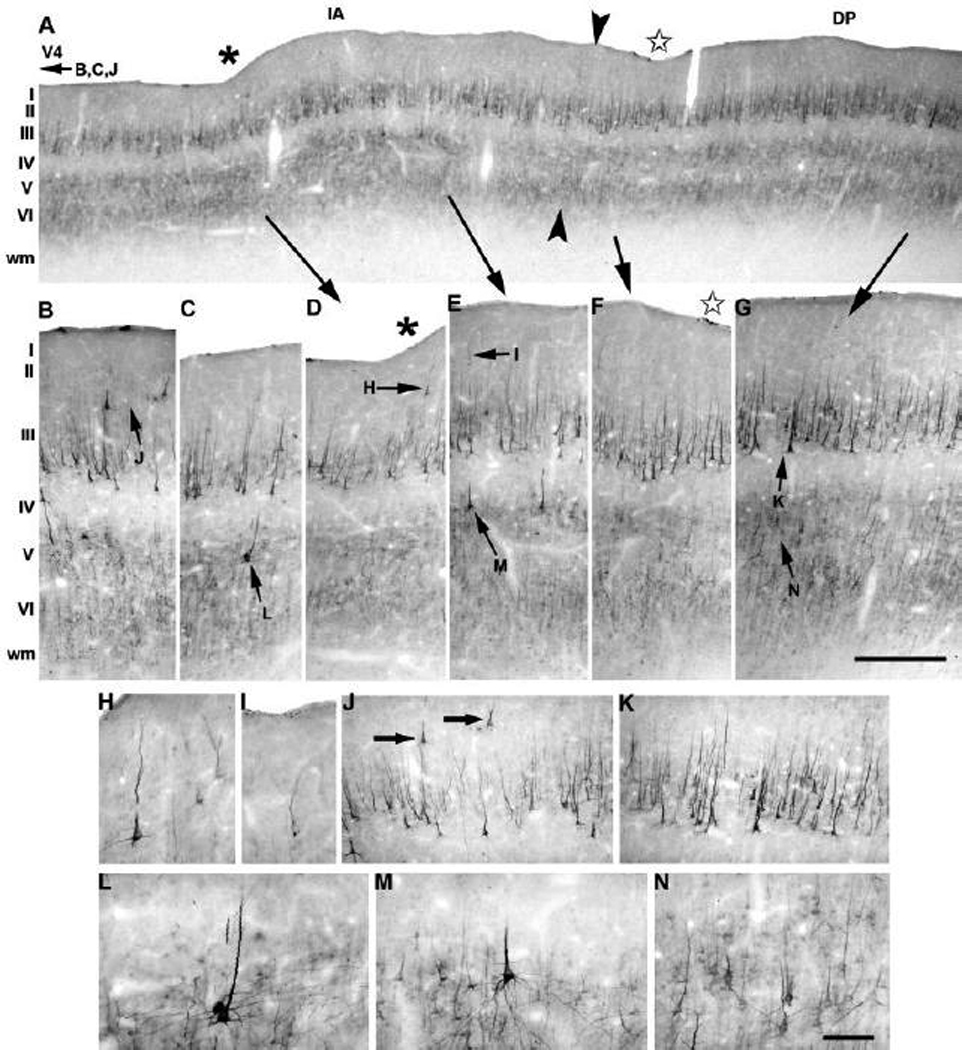
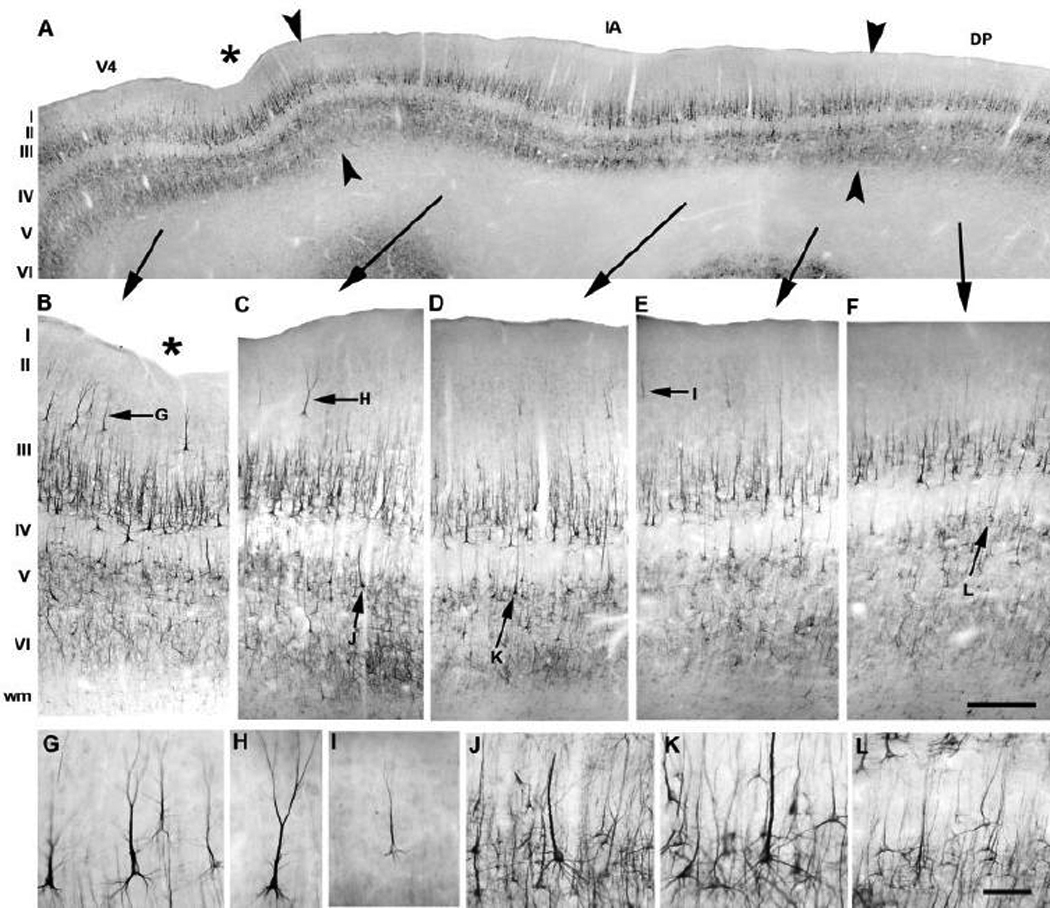
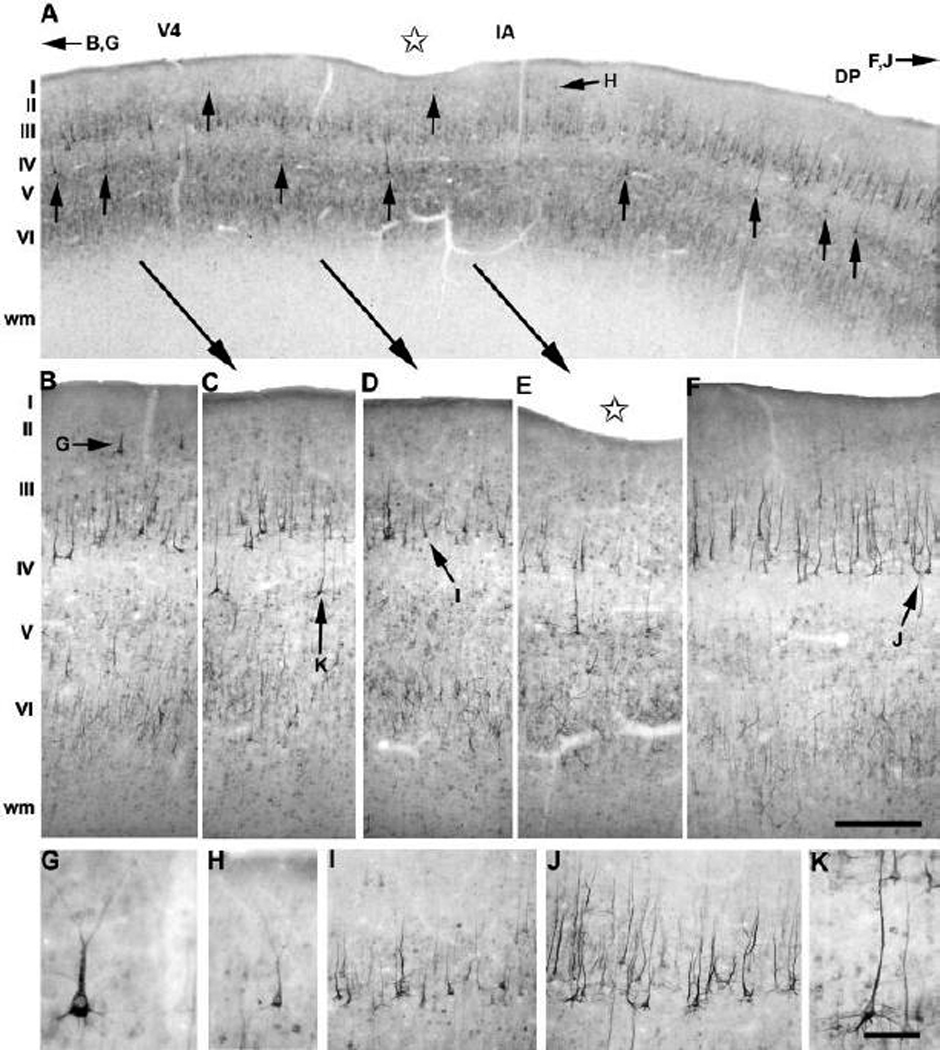
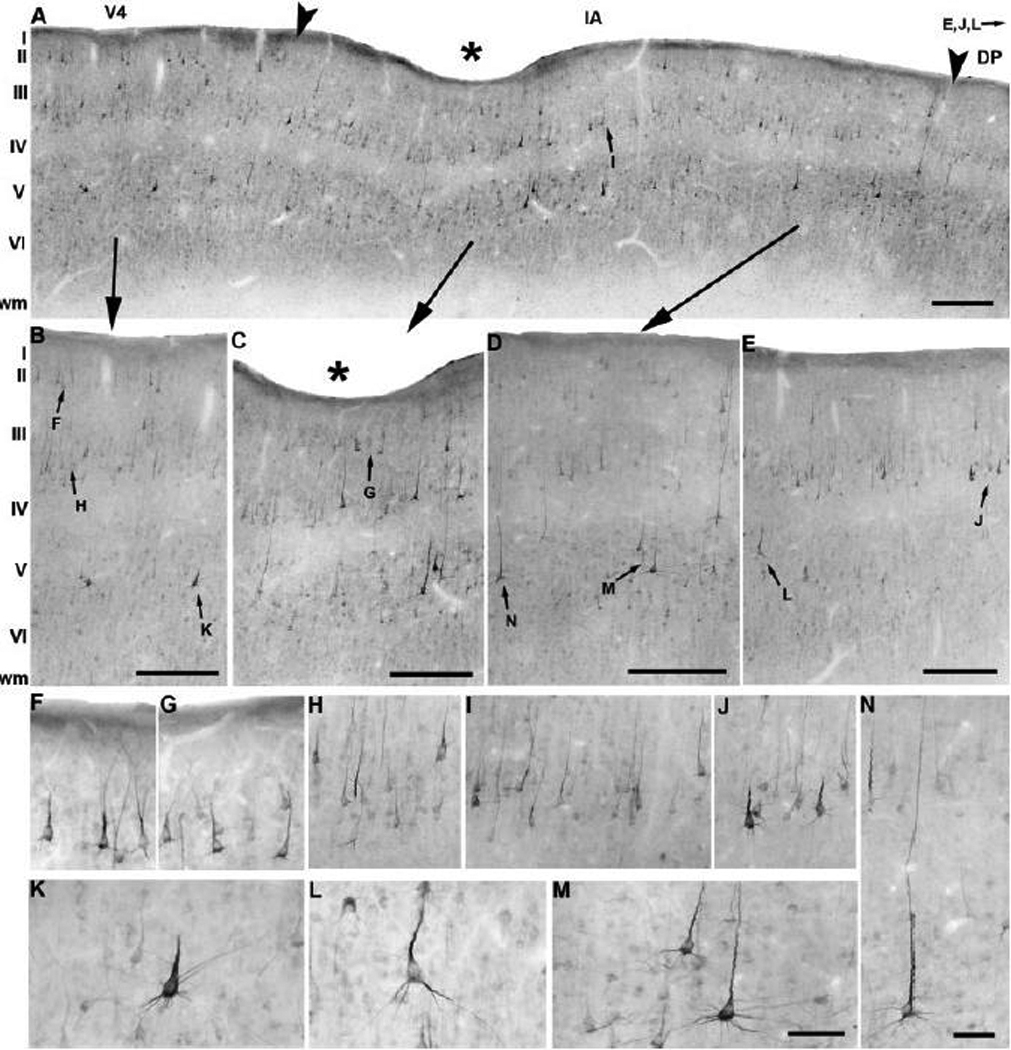
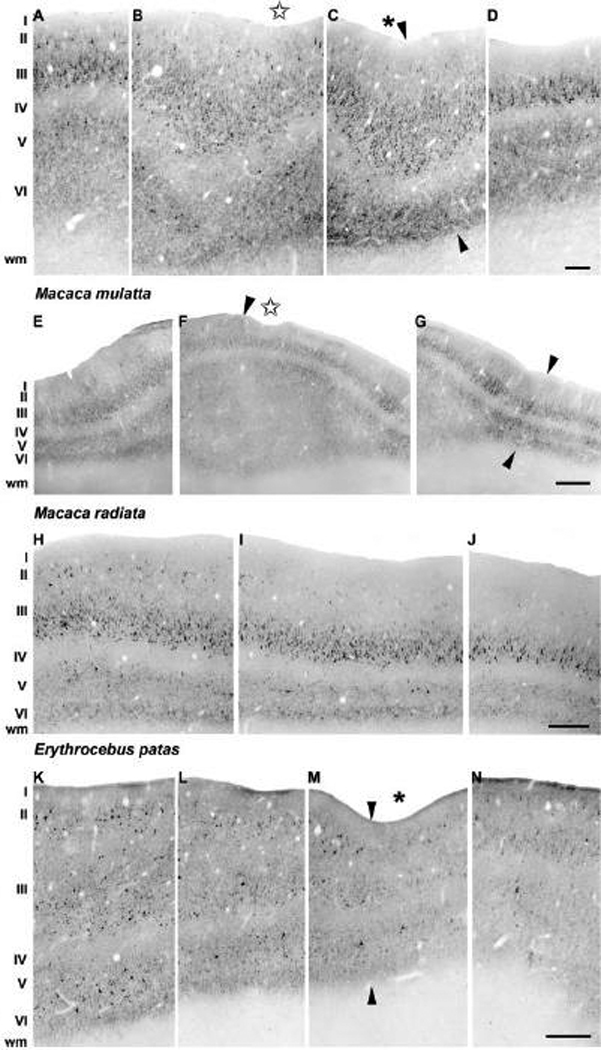
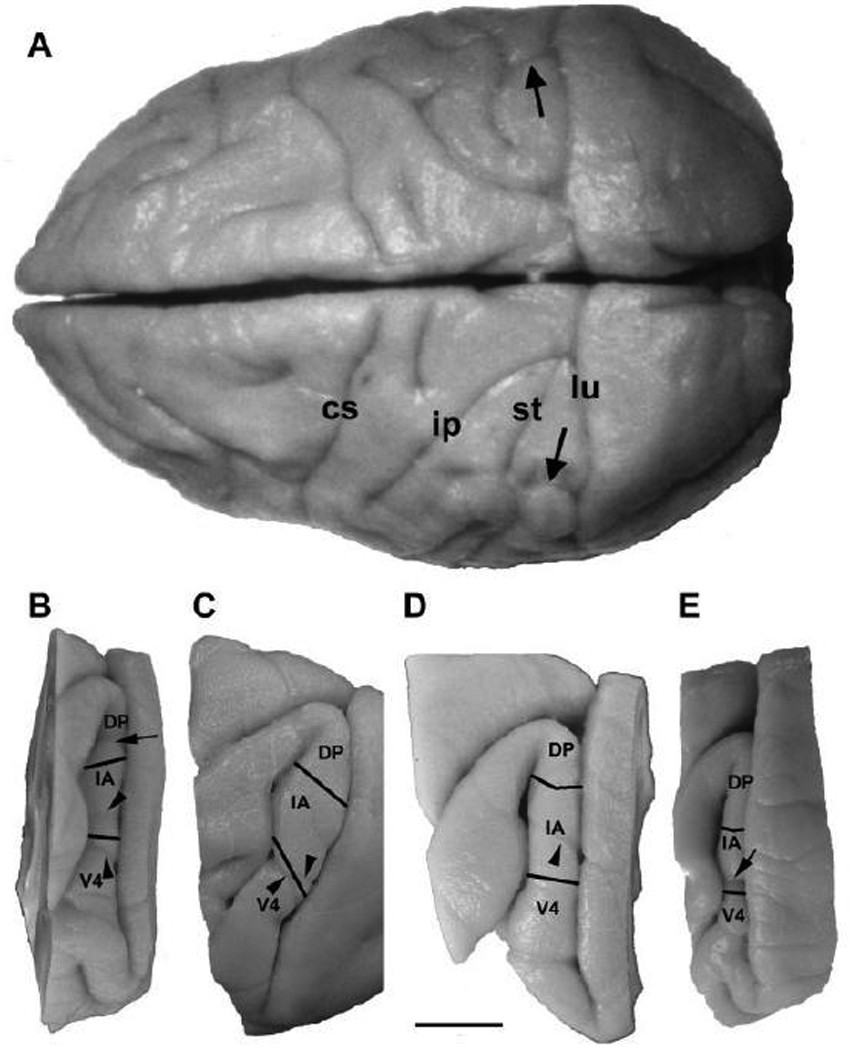
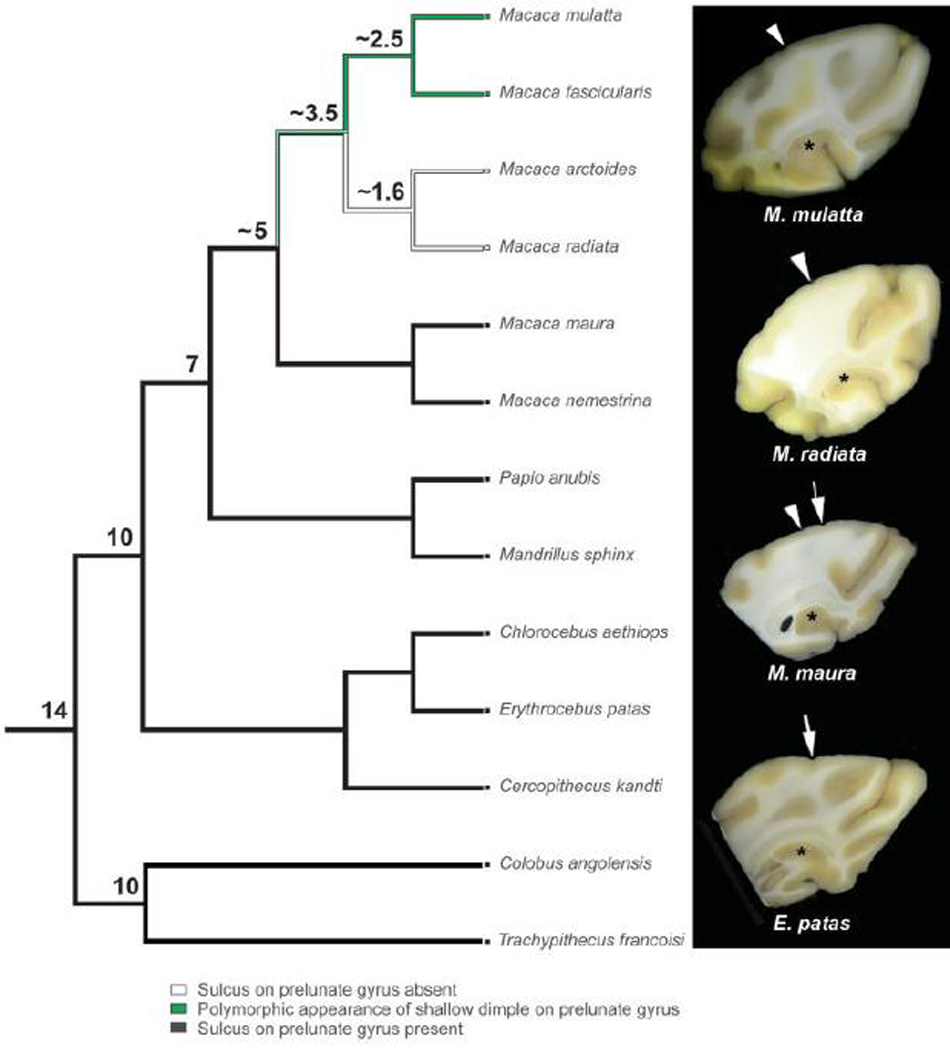
Anatomical and electrophysiological studies have revealed a complex organization in the macaque prelunate gyrus. We investigated the morphology and architecture of the prelunate gyrus in Old World monkeys. In Macaca nemestrina, we observed a sulcus crossing the prelunate gyrus within 2 mm of the vertical meridian representation. In other macaque species and other cercopithecines, we observed substantial variations in sulcal morphology across the prelunate gyrus. We did not find a sulcus in all species, and the location and depth of that indentation on the gyrus varied among species. A deep sulcus was observed in all species that emerged earlier in evolution than macaques, such as guenons, baboons, and colobines. We analyzed the regional and parcellation features of the prelunate gyrus in three macaque species, M. maura, M. mulatta, and M. radiata, and in Erythrocebus patas, with emphasis on the relation of structure to the distribution of prelunate visual areas. Nonphosphorylated neurofilament protein immunoreactivity permitted the delineation of a novel area in the prelunate gyrus of Old World monkeys, located around the prelunate sulcus. Species-specific patterns were also observed in the prelunate gyrus of the patas monkey compared to macaques. These observations, as well as a cladistic analysis of the data, suggest an expanded and diversified organization of the prelunate gyrus in some cercopithecoids that may reflect adaptation to specific ecological environments. It was, however, progressively lost in most macaques, being retained only in species that diverged early in the evolution of the genus Macaca, such as M. nemestrina and M. maura.
Figures
References
-
- Allman JM, Kaas JH. A crescent-shaped cortical visual area surrounding the middle temporal area (MT) in the owl monkey (Aotus trivirgatus) Brain Res. 1974a;81:199–213. - PubMed
-
- Allman JM, Kaas JH. The organization of the second visual area (V II) in the owl monkey: a second order transformation of the visual hemifield. Brain Res. 1974b;76:247–265. - PubMed
-
- Allman JM, Kaas JH. The dorsomedial cortical visual area: a third tier area in the occipital lobe of the owl monkey (Aotus trivirgatus) Brain Res. 1975;100:473–487. - PubMed
-
- Allman JM, Kaas JH. Representation of the visual field on the medical wall of occipital-parietal cortex in the owl monkey. Science. 1976;191:572–575. - PubMed
-
- Allman JM, Kaas JH, Lane RH. The middle temporal visual area (MT) in the bushbaby, Galago senegalensis. Brain Res. 1973;57:197–202. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
