

Microstimulation of the somatosensory cortex can substitute for vibrissa stimulation during Pavlovian conditioning
- PMID: 16782811
- PMCID: PMC1479767
- DOI: 10.1073/pnas.0603584103
Microstimulation of the somatosensory cortex can substitute for vibrissa stimulation during Pavlovian conditioning
Abstract
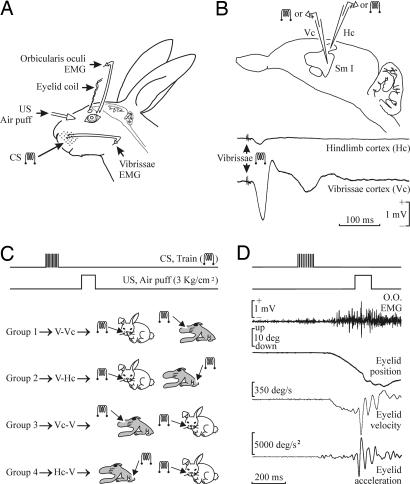
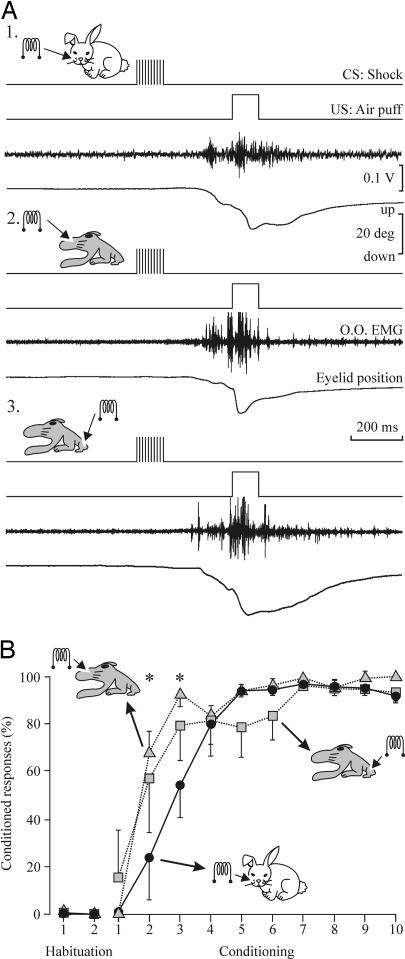
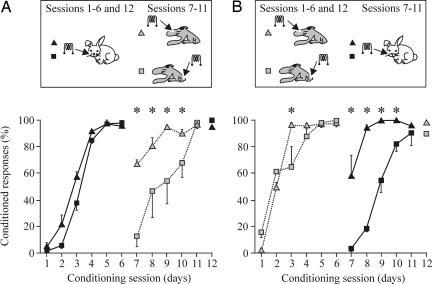
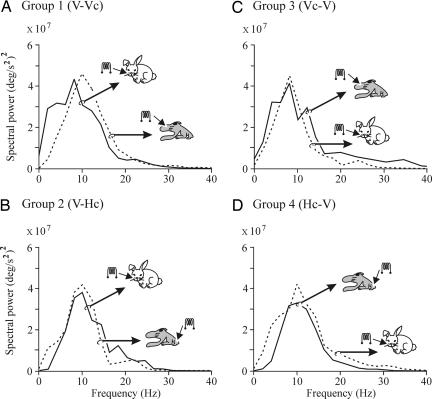
The primary somatosensory cortex (S1) contains a map representation of the body surface. We hypothesized that S1 stimulation can successfully substitute for (or be substituted by) direct stimulation of skin receptors. We prepared rabbits for evoking eyelid conditioned responses (CRs) using a trace "shock-air puff" paradigm. In a first series of experiments, animals received a conditioned stimulus (CS, a train of electrical pulses) in the whisker pad or in the S1 areas for vibrissae or for the hind limb. In the three cases, the CS was followed 250 ms from its end by an air puff presented to the cornea as an unconditioned stimulus (US). Learning curves from the three groups presented similar values, although animals stimulated with a central CS acquired their CRs faster. In a second series of experiments, animals were divided into four groups and were presented either centrally or peripherally with the same CS for six conditioning sessions. Then, the CS was switched from central to peripheral, or vice versa, for 5 additional days. Conditioned animals were not able to discriminate between peripheral (vibrissae) stimuli and stimuli presented to the corresponding S1 (vibrissae) area, but they were able to discriminate between CSs presented to S1 (hind limb) and body (vibrissae) regions. The kinetic properties of evoked CRs were not modified by CS switching. It is proposed that S1 allows the construction of somatosensory percepts of the body surface but does not allow distinguishing the central or peripheral location of the evoking stimuli.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures

References
-
- Lafuente V. de, Romo R. Nat. Neurosci. 2005;8:1698–1703. - PubMed
-
- Romo R., Salinas E. Annu. Rev. Neurosci. 2001;24:107–137. - PubMed
-
- Wilent W. B., Contreras D. Nat. Neurosci. 2005;8:1364–1370. - PubMed
-
- Romo R., Hernández A., Zainos A., Salinas E. Nature. 1998;392:387–390. - PubMed
-
- Salzman C. D., Britten K. H., Newsome W. T. Nature. 1990;346:174–177. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
