

FAT10/diubiquitin-like protein-deficient mice exhibit minimal phenotypic differences
- PMID: 16782901
- PMCID: PMC1489174
- DOI: 10.1128/MCB.00966-05
FAT10/diubiquitin-like protein-deficient mice exhibit minimal phenotypic differences
Abstract
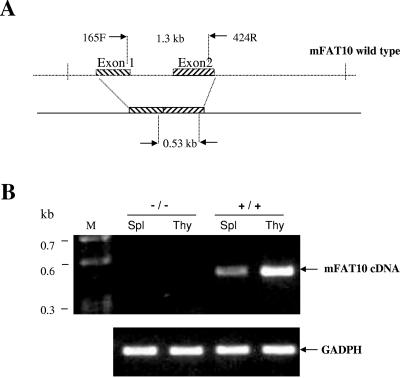
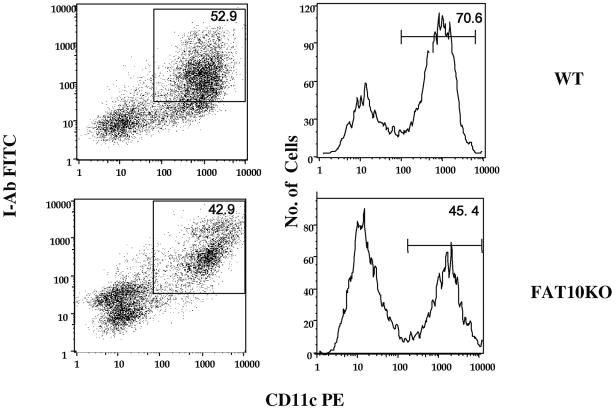
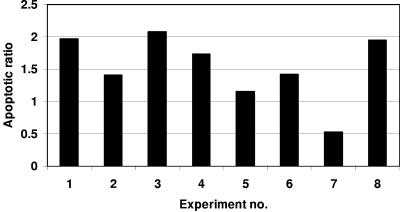
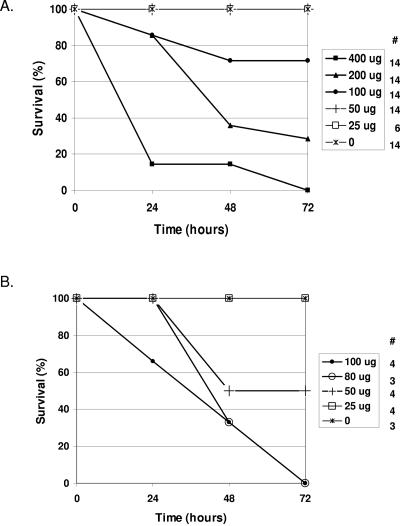
The FAT10 gene encodes a diubiquitin-like protein containing two tandem head-to-tail ubiquitin-like domains. There is a high degree of similarity between murine and human FAT10 sequences at both the mRNA and protein levels. In various cell lines, FAT10 expression was shown to be induced by gamma interferon or by tumor necrosis factor alpha. In addition, FAT10 expression was found to be up-regulated in some Epstein-Barr virus-infected B-cell lines, in activated dendritic cells, and in several epithelial tumors. However, forced expression of FAT10 in cultured cells was also found to produce apoptotic cell death. Overall, these findings suggest that FAT10 may modulate cellular growth or cellular viability. Here we describe the steps to generate, by genetic targeting, a FAT10 gene knockout mouse model. The FAT10 knockout homozygous mice are viable and fertile. No gross lesions or obvious histological differences were found in these mutated mice. Examination of lymphocyte populations from spleen, thymus, and bone marrow did not reveal any abnormalities. However, flow cytometry analysis demonstrated that the lymphocytes of FAT10 knockout mice were, on average, more prone to spontaneous apoptotic death. Physiologically, these mice demonstrated a high level of sensitivity toward endotoxin challenge. These findings indicate that FAT10 may function as a survival factor.
Figures


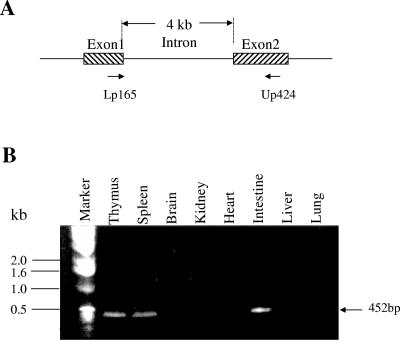
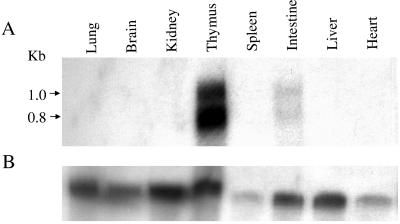
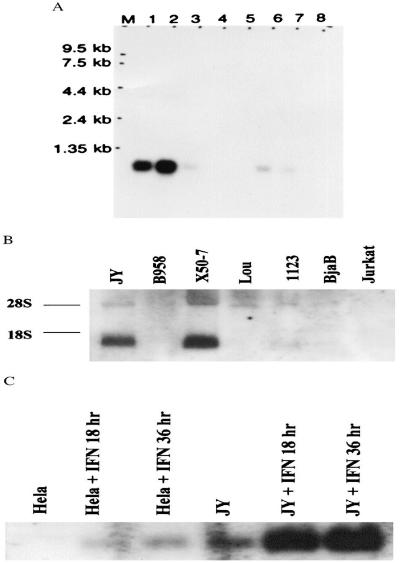
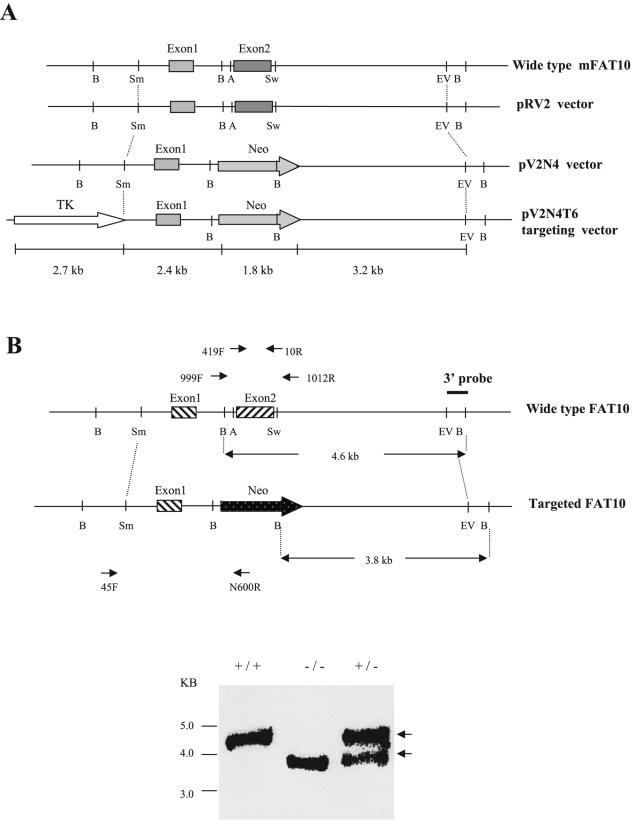
References
-
- Bates, E. E., O. Ravel, M. C. Dieu, S. Ho, C. Guret, J. M. Bridon, S. Ait-Yahia, F. Briere, C. Caux, J. Banchereau, and S. Lebecque. 1997. Identification and analysis of a novel member of the ubiquitin family expressed in dendritic cells and mature B cells. Eur. J. Immunol. 27:2471-2477. - PubMed
-
- Boddy, M. N., K. Howe, L. D. Etkin, E. Solomon, and P. S. Freemont. 1996. PIC 1, a novel ubiquitin-like protein which interacts with the PML component of a multiprotein complex that is disrupted in acute promyelocytic leukaemia. Oncogene 13:971-982. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases