

Phosphoinositide-dependent kinase 1 targets protein kinase A in a pathway that regulates interleukin 4
- PMID: 16785309
- PMCID: PMC2118337
- DOI: 10.1084/jem.20051715
Phosphoinositide-dependent kinase 1 targets protein kinase A in a pathway that regulates interleukin 4
Abstract
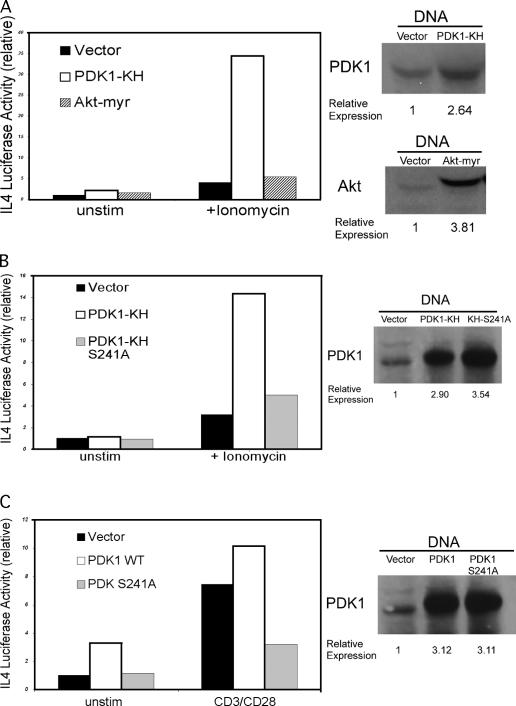
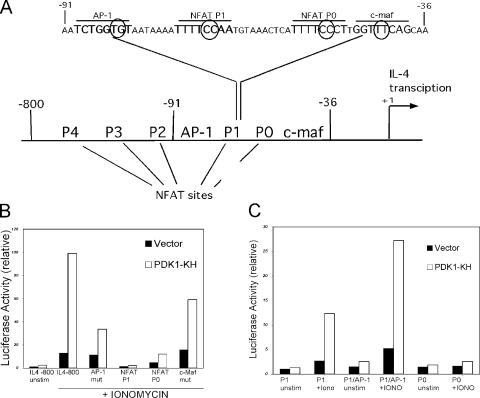
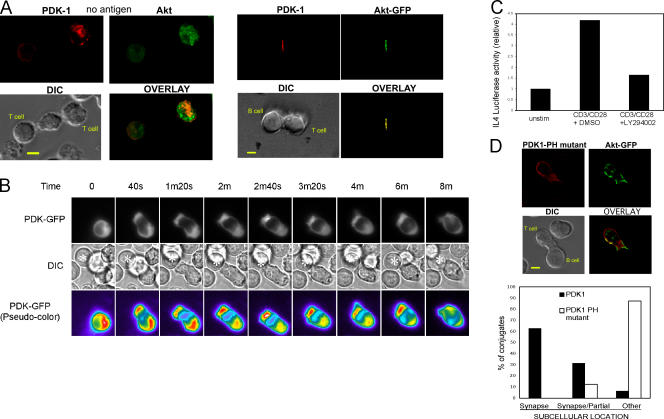
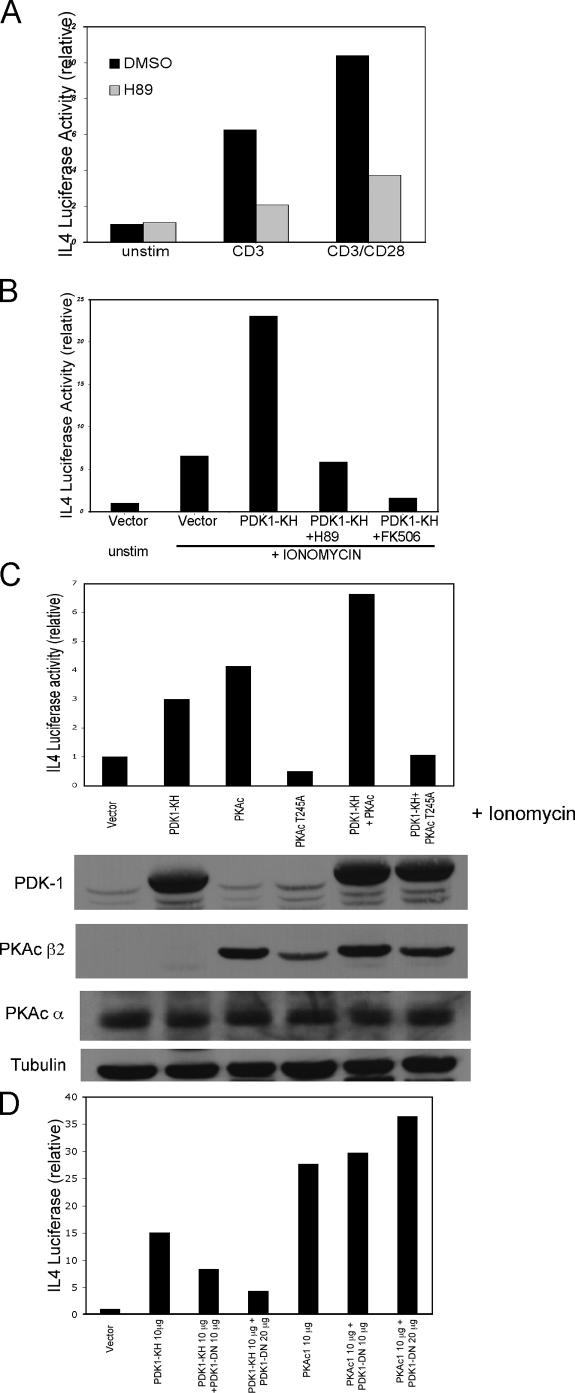
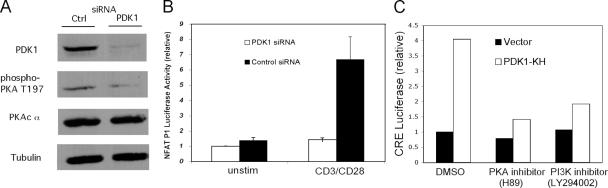
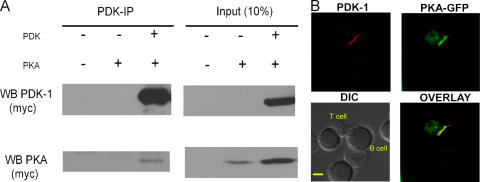
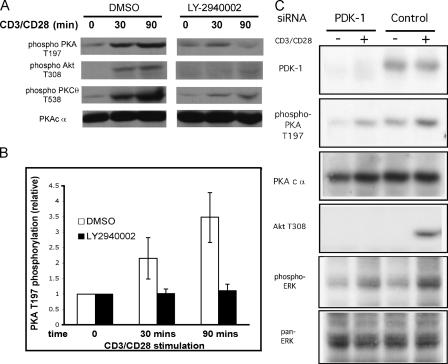
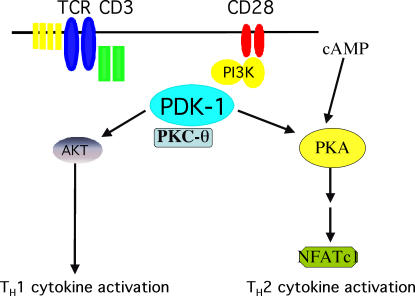
CD28 plays a critical role in T cell immune responses. Although the kinase Akt has been shown to act downstream of CD28 in T helper (Th)1 cytokine induction, it does not induce Th2 cytokines such as interleukin 4 (IL-4). We recently reported that phosphoinositide-dependent kinase 1 (PDK1) partially corrects the defect in IL-4 production present in CD28-deficient T cells, suggesting that PDK1 regulates IL-4 independently of Akt. We now describe a signaling pathway in which PDK1 targets IL-4 in the murine Th2 cell line D10. PDK1-mediated activation of this pathway is dependent on protein kinase A (PKA) and the nuclear factor of activated T cells (NFAT) P1 transcriptional element in the IL-4 promoter. PDK1 localizes to the immune synapse in a phosphatidylinositol 3-kinase-dependent manner, partially colocalizes with PKA at the synapse, and physically interacts with PKA. In RNA interference knockdown experiments, PDK1 is necessary for phosphorylation of PKA in T cells, as well as for activation of the IL-4 NFAT P1 element by the T cell receptor (TCR) and CD28. Phosphorylation of the critical PKA threonine residue is stimulated by engagement of TCR/CD28 via a PDK1-dependent mechanism. These findings together define a pathway linking the kinases PDK1 and PKA in the induction of the Th2 cytokine IL-4.
Figures
References
-
- Salomon, B., and J.A. Bluestone. 2001. Complexities of CD28/B7: CTLA-4 costimulatory pathways in autoimmunity and transplantation. Annu. Rev. Immunol. 19:225–252. - PubMed
-
- Fraser, J.D., B.A. Irving, G.R. Crabtree, and A. Weiss. 1991. Regulation of interleukin-2 gene enhancer activity by the T cell accessory molecule CD28. Science. 251:313–316. - PubMed
-
- Rulifson, I.C., A.I. Sperling, P.E. Fields, F.W. Fitch, and J.A. Bluestone. 1997. CD28 costimulation promotes the production of Th2 cytokines. J. Immunol. 158:658–665. - PubMed
-
- Shahinian, A., K. Pfeffer, K.P. Lee, T.M. Kundig, K. Kishihara, A. Wakeham, K. Kawai, P.S. Ohashi, C.B. Thompson, and T.W. Mak. 1993. Differential T cell costimulatory requirements in CD28-deficient mice. Science. 261:609–612. - PubMed
-
- Kane, L.P., P.G. Andres, K.C. Howland, A.K. Abbas, and A. Weiss. 2001. Akt provides the CD28 costimulatory signal for up-regulation of IL-2 and IFN-γ but not TH2 cytokines. Nat. Immunol. 2:37–44. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
