

Molecular cloning of a Trypanosoma cruzi cell surface casein kinase II substrate, Tc-1, involved in cellular infection
- PMID: 16790765
- PMCID: PMC1489715
- DOI: 10.1128/IAI.00045-06
Molecular cloning of a Trypanosoma cruzi cell surface casein kinase II substrate, Tc-1, involved in cellular infection
Abstract
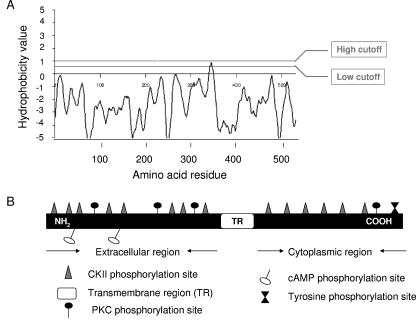
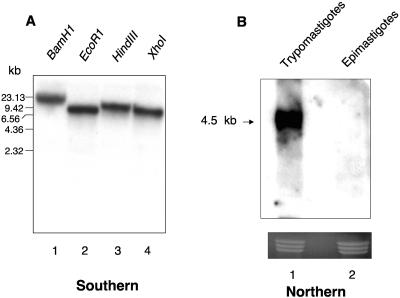
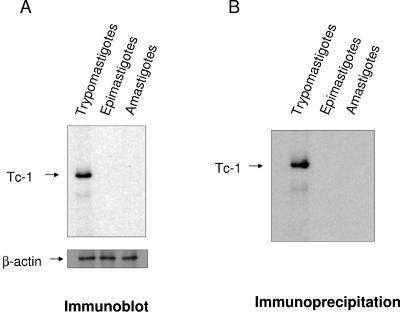
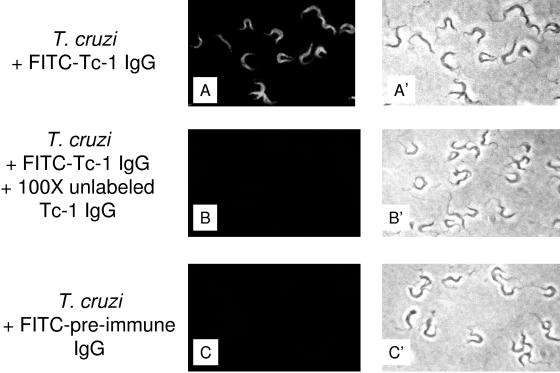
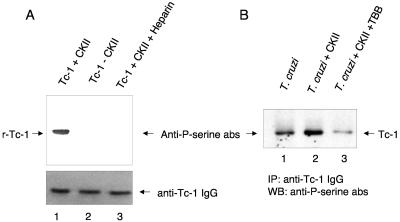
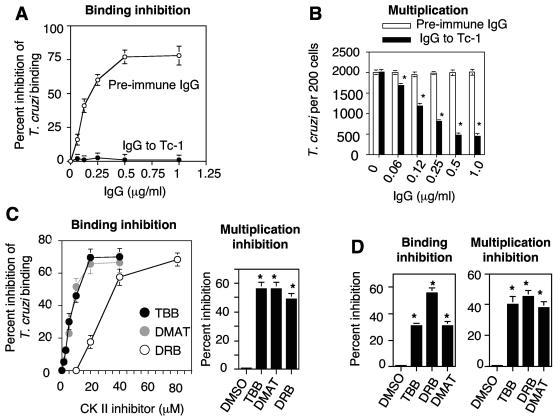
In this work, we report the cloning and characterization of the first cell surface casein kinase II (CKII) substrate (Tc-1) of Trypanosoma cruzi, the causative agent of Chagas' disease. Analysis of the gene sequence revealed a 1,653-bp open reading frame coding for 550 amino acid residues. Northern blot analysis showed a 4.5-kb transcript that is expressed in invasive trypomastigotes but not in noninvasive epimastigote forms of T. cruzi. Southern blot analysis indicates that Tc-1 is a single-copy gene. At the amino acid level, Tc-1 displayed 95% and 99% identity to two hypothetical proteins recently reported by the T. cruzi genome project. Analysis of the translated amino acid sequence indicates that the Tc-1 gene has a putative transmembrane domain with multiple cytoplasmic and extracellular CKII phosphosites. Exogenous human CKII was able to phosphorylate serine residues on both recombinant Tc-1 and Tc-1 of intact trypomastigotes. This phosphorylation was inhibited by the CKII inhibitors heparin and 4,5,6,7,-tetrabromo-2-azabenzimidazole. Immunoblots of solubilized trypomastigotes, epimastigotes, and amastigotes probed with anti-recombinant Tc-1 immunoglobulin G revealed a 62-kDa protein that is expressed only in infective trypomastigotes. Immunoprecipitation of labeled surface proteins of trypomastigotes indicated that the 62-kDa protein is a surface protein, and we found that the protein is uniformly distributed on the surface of trypomastigotes by direct immunofluorescence. Antibodies to Tc-1 effectively blocked trypomastigote invasion of host cells and consequently reduced parasite load. Preincubation of either trypomastigotes or myoblasts with CKII inhibitors blocked T. cruzi infection. Thus, for the first time, we describe a cell surface CKII substrate of a protozoan parasite that is phosphorylated by human CKII and that is involved in cellular infection.
Figures

Similar articles
-
Molecular Characterization of Trypanosoma cruzi Tc8.2 Gene Indicates Two Differential Locations for the Encoded Protein in Epimastigote and Trypomastigote Forms.Korean J Parasitol. 2015 Aug;53(4):483-8. doi: 10.3347/kjp.2015.53.4.483. Epub 2015 Aug 25. Korean J Parasitol. 2015. PMID: 26323848 Free PMC article.
-
Trypanosoma cruzi: characterization of two recombinant antigens with potential application in the diagnosis of Chagas' disease.Exp Parasitol. 1993 Feb;76(1):1-12. doi: 10.1006/expr.1993.1001. Exp Parasitol. 1993. PMID: 8467895
-
Phospholipase A1: a novel virulence factor in Trypanosoma cruzi.Mol Biochem Parasitol. 2013 Feb;187(2):77-86. doi: 10.1016/j.molbiopara.2012.12.004. Epub 2012 Dec 27. Mol Biochem Parasitol. 2013. PMID: 23275096
-
Molecular mimicry between Trypanosoma cruzi and host nervous tissues.Acta Cient Venez. 1992;43(6):330-40. Acta Cient Venez. 1992. PMID: 1343745 Review.
-
Proteins anchored via glycosylphosphatidylinositol and solubilizing phospholipases in Trypanosoma cruzi.Biol Res. 1993;26(1-2):285-312. Biol Res. 1993. PMID: 7670541 Review.
Cited by
-
Perspectives on the Trypanosoma cruzi-host cell receptor interactions.Parasitol Res. 2009 Jun;104(6):1251-60. doi: 10.1007/s00436-009-1383-3. Epub 2009 Mar 13. Parasitol Res. 2009. PMID: 19283409 Free PMC article. Review.
-
Stress induces changes in the phosphorylation of Trypanosoma cruzi RNA polymerase II, affecting its association with chromatin and RNA processing.Eukaryot Cell. 2014 Jul;13(7):855-65. doi: 10.1128/EC.00066-14. Epub 2014 May 9. Eukaryot Cell. 2014. PMID: 24813189 Free PMC article.
-
Molecular analysis of early host cell infection by Trypanosoma cruzi.Front Biosci. 2008 May 1;13:3714-34. doi: 10.2741/2961. Front Biosci. 2008. PMID: 18508467 Free PMC article. Review.
-
VNI cures acute and chronic experimental Chagas disease.J Infect Dis. 2013 Aug 1;208(3):504-11. doi: 10.1093/infdis/jit042. Epub 2013 Jan 31. J Infect Dis. 2013. PMID: 23372180 Free PMC article.
-
Protein kinase CK2: a potential therapeutic target for diverse human diseases.Signal Transduct Target Ther. 2021 May 17;6(1):183. doi: 10.1038/s41392-021-00567-7. Signal Transduct Target Ther. 2021. PMID: 33994545 Free PMC article. Review.
References
-
- Burleigh, B. B., and A. M. Woolsey. 2002. Cell signaling and Trypanosoma cruzi invasion. Cell. Microbiol. 4:701-711. - PubMed
-
- El-Sayed, N. M., P. J. Myler, G. Blandin, M. Berriman, J. Crabtree, G. Aggarwal, E. Caler, H. Renauld, E. A. Worthey, C. Hertz-Fowler, E. Ghedin, C. Peacock, D. C. Bartholomeu, B. J. Haas, A. N. Tran, J. R. Wortman, U. C. Alsmark, S. Angiuoli, A. Anupama, J. Badger, F. Bringaud, E. Cadag, J. M. Carlton, G. C. Cerqueira, T. Creasy, A. L. Delcher, A. Djikeng, T. M. Embley, C. Hauser, A. C. Ivens, S. K. Kummerfeld, J. B. Pereira-Leal, D. Nilsson, J. Peterson, S. L. Salzberg, J. Shallom, J. C. Silva, J. Sundaram, S. Westenberger, O. White, S. E. Melville, J. E. Donelson, B. Andersson, K. D. Stuart, and N. Hall. 2005. Comparative genomics of trypanosomatid parasitic protozoa. Science 309:404-409. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- T32 HL007737/HL/NHLBI NIH HHS/United States
- MD 000104/MD/NIMHD NIH HHS/United States
- HL 007737/HL/NHLBI NIH HHS/United States
- G12 RR003032/RR/NCRR NIH HHS/United States
- GM 08037/GM/NIGMS NIH HHS/United States
- RR 003032/RR/NCRR NIH HHS/United States
- S21 MD000104/MD/NIMHD NIH HHS/United States
- S06 GM008037/GM/NIGMS NIH HHS/United States
- T32 AI007281/AI/NIAID NIH HHS/United States
- F31 AI056667/AI/NIAID NIH HHS/United States
- AI 056667/AI/NIAID NIH HHS/United States
- GM 05994/GM/NIGMS NIH HHS/United States
- AI 07281/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
