

The type III pseudomonal exotoxin U activates the c-Jun NH2-terminal kinase pathway and increases human epithelial interleukin-8 production
- PMID: 16790784
- PMCID: PMC1489742
- DOI: 10.1128/IAI.02045-05
The type III pseudomonal exotoxin U activates the c-Jun NH2-terminal kinase pathway and increases human epithelial interleukin-8 production
Abstract
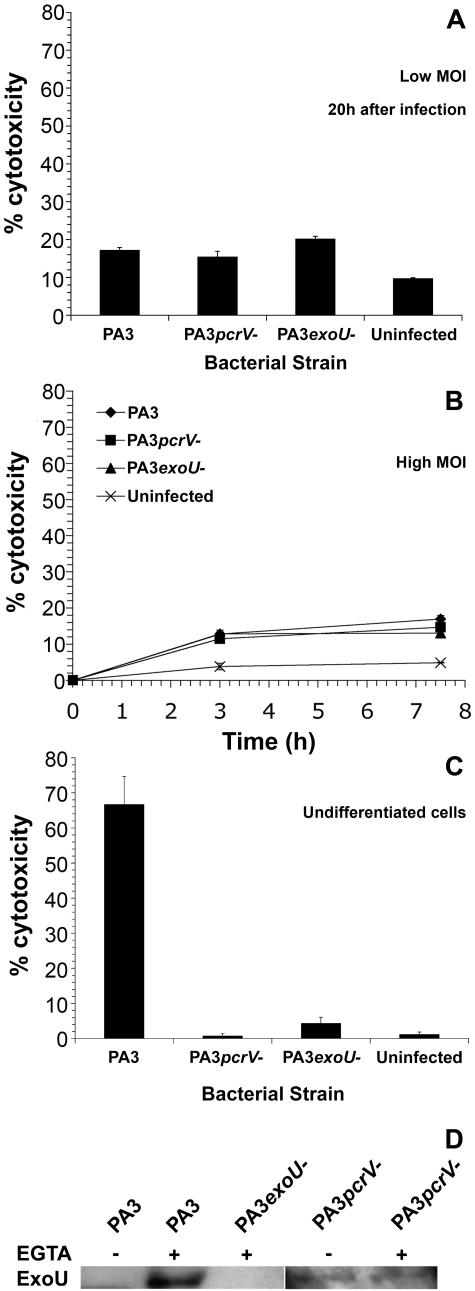
Microbial interactions with host cell signaling pathways are key determinants of the host cell response to infection. Many toxins secreted by bacterial type III secretion systems either stimulate or inhibit the host inflammatory response. We investigated the role of type III secreted toxins of the lung pathogen Pseudomonas aeruginosa in the inflammatory response of human respiratory epithelial cells to infection. Using bacteria with specific gene deletions, we found that interleukin-8 production by these cells was almost entirely dependent on bacterial type III secretion of exotoxin U (ExoU), a phospholipase, although other bacterial factors are involved. ExoU activated the c-Jun NH(2)-terminal kinase pathway, stimulating the phosphorylation and activation of mitogen-activated kinase kinase 4, c-Jun NH(2)-terminal kinase, and c-Jun. This in turn increased levels of transcriptionally competent activator protein-1. Although this pathway was dependent on the lipase activity of ExoU, it was independent of cell death. Activation of mitogen-activated kinase signaling by ExoU in this fashion is a novel mechanism by which a bacterial product can initiate a host inflammatory response, and it may result in increased epithelial permeability and bacterial spread.
Figures






Similar articles
-
Effector ExoU from the type III secretion system is an important modulator of gene expression in lung epithelial cells in response to Pseudomonas aeruginosa infection.Infect Immun. 2003 Oct;71(10):6035-44. doi: 10.1128/IAI.71.10.6035-6044.2003. Infect Immun. 2003. PMID: 14500525 Free PMC article.
-
Nintedanib modulates surfactant protein-D expression in A549 human lung epithelial cells via the c-Jun N-terminal kinase-activator protein-1 pathway.Pulm Pharmacol Ther. 2015 Jun;32:29-36. doi: 10.1016/j.pupt.2015.03.001. Epub 2015 Apr 2. Pulm Pharmacol Ther. 2015. PMID: 25843005
-
Activation of extracellular signal-regulated kinase and c-Jun-NH(2)-terminal kinase but not p38 mitogen-activated protein kinases is required for RRR-alpha-tocopheryl succinate-induced apoptosis of human breast cancer cells.Cancer Res. 2001 Sep 1;61(17):6569-76. Cancer Res. 2001. PMID: 11522656
-
Inflammatory role of AMP-activated protein kinase signaling in an experimental model of toxic smoke inhalation injury.Crit Care Med. 2013 Jan;41(1):120-32. doi: 10.1097/CCM.0b013e318265f653. Crit Care Med. 2013. PMID: 23222262
-
1alpha,25-dihydroxyvitamin D3 stimulates activator protein 1 DNA-binding activity by a phosphatidylinositol 3-kinase/Ras/MEK/extracellular signal regulated kinase 1/2 and c-Jun N-terminal kinase 1-dependent increase in c-Fos, Fra1, and c-Jun expression in human keratinocytes.J Invest Dermatol. 2003 Apr;120(4):561-70. doi: 10.1046/j.1523-1747.2003.12095.x. J Invest Dermatol. 2003. PMID: 12648218
Cited by
-
Bacterial Sphingomyelinases and Phospholipases as Virulence Factors.Microbiol Mol Biol Rev. 2016 Jun 15;80(3):597-628. doi: 10.1128/MMBR.00082-15. Print 2016 Sep. Microbiol Mol Biol Rev. 2016. PMID: 27307578 Free PMC article. Review.
-
ExoU activates NF-κB and increases IL-8/KC secretion during Pseudomonas aeruginosa infection.PLoS One. 2012;7(7):e41772. doi: 10.1371/journal.pone.0041772. Epub 2012 Jul 26. PLoS One. 2012. PMID: 22848596 Free PMC article.
-
Bacterial Nucleotidyl Cyclase Inhibits the Host Innate Immune Response by Suppressing TAK1 Activation.Infect Immun. 2017 Aug 18;85(9):e00239-17. doi: 10.1128/IAI.00239-17. Print 2017 Sep. Infect Immun. 2017. PMID: 28652310 Free PMC article.
-
Pseudomonas aeruginosa Toxin ExoU as a Therapeutic Target in the Treatment of Bacterial Infections.Microorganisms. 2019 Dec 16;7(12):707. doi: 10.3390/microorganisms7120707. Microorganisms. 2019. PMID: 31888268 Free PMC article. Review.
-
Multiple roles of phospholipase A2 during lung infection and inflammation.Infect Immun. 2008 Jun;76(6):2259-72. doi: 10.1128/IAI.00059-08. Epub 2008 Apr 14. Infect Immun. 2008. PMID: 18411286 Free PMC article. Review. No abstract available.
References
-
- Bishara, J., L. Leibovici, S. Ashkenazi, Z. Samra, and S. Pitlik. 2000. Seven-year study of bacteraemic pneumonia in a single institution. Eur. J. Clin. Microbiol. Infect. Dis. 19:926-931. - PubMed
-
- Bogoyevitch, M. A., I. Boehm, A. Oakley, A. J. Ketterman, and R. K. Barr. 2004. Targeting the JNK MAPK cascade for inhibition: basic science and therapeutic potential. Biochim. Biophys. Acta 1697:89-101. - PubMed
-
- Bonfield, T. L., J. R. Panuska, M. W. Konstan, K. A. Hilliard, J. B. Hilliard, H. Ghnaim, and M. Berger. 1995. Inflammatory cytokines in cystic fibrosis lungs. Am. J. Respir. Crit. Care Med. 152:2111-2118. - PubMed
-
- Chastre, J., and J. Y. Fagon. 2002. Ventilator-associated pneumonia. Am. J. Respir. Crit. Care Med. 165:867-903. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
