

Sustained elevation of dendritic calcium evokes widespread endocannabinoid release and suppression of synapses onto cerebellar Purkinje cells
- PMID: 16793891
- PMCID: PMC6673843
- DOI: 10.1523/JNEUROSCI.1280-06.2006
Sustained elevation of dendritic calcium evokes widespread endocannabinoid release and suppression of synapses onto cerebellar Purkinje cells
Abstract
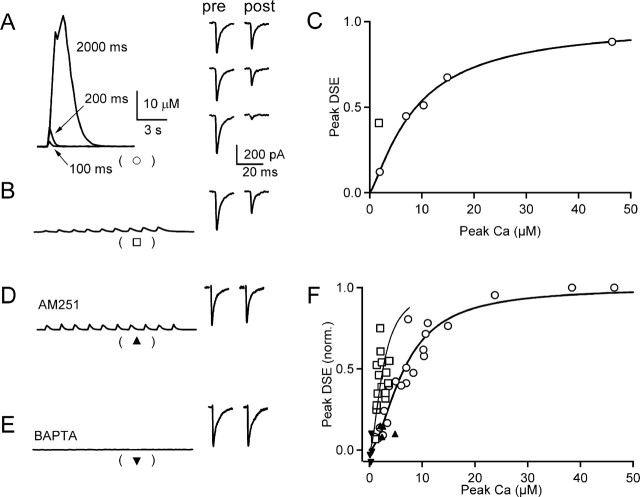
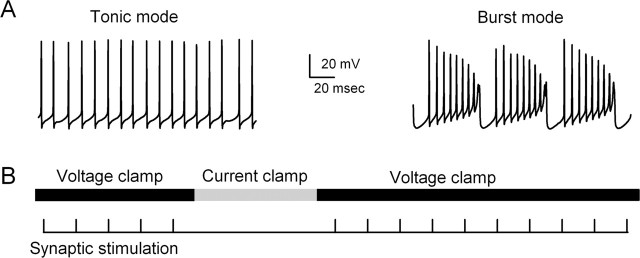
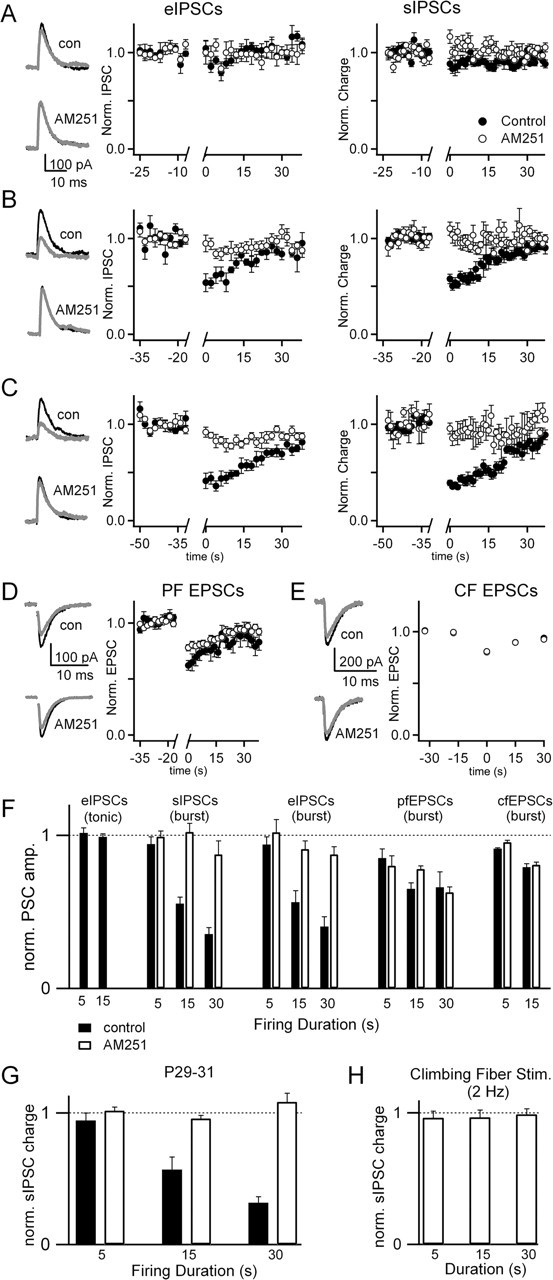
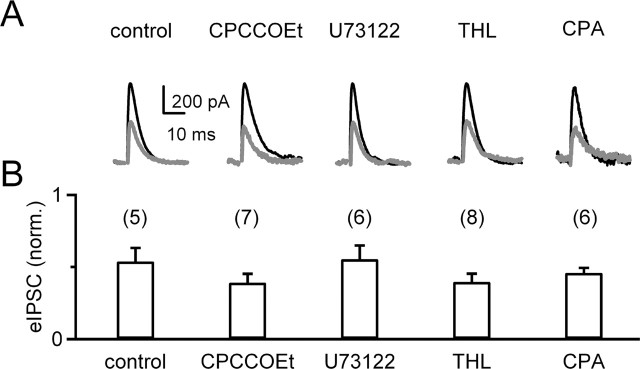
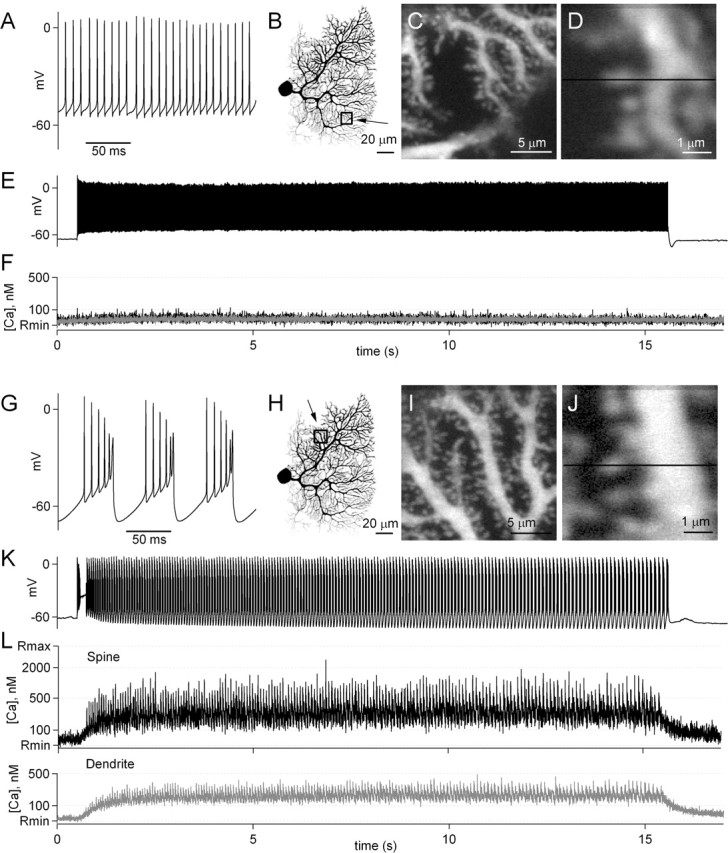
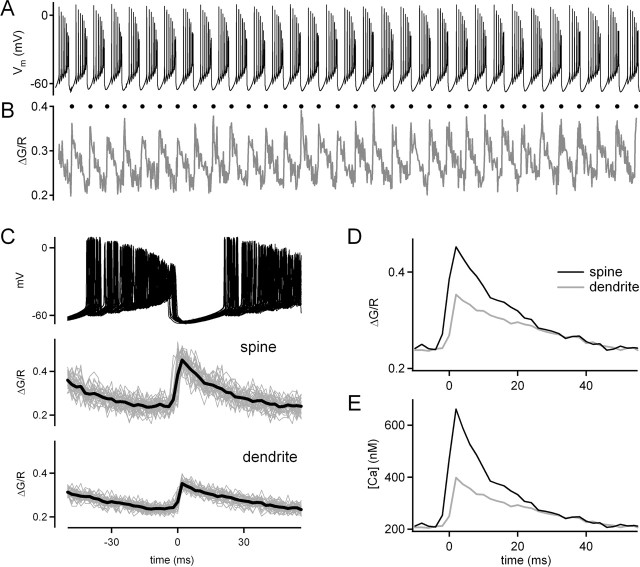
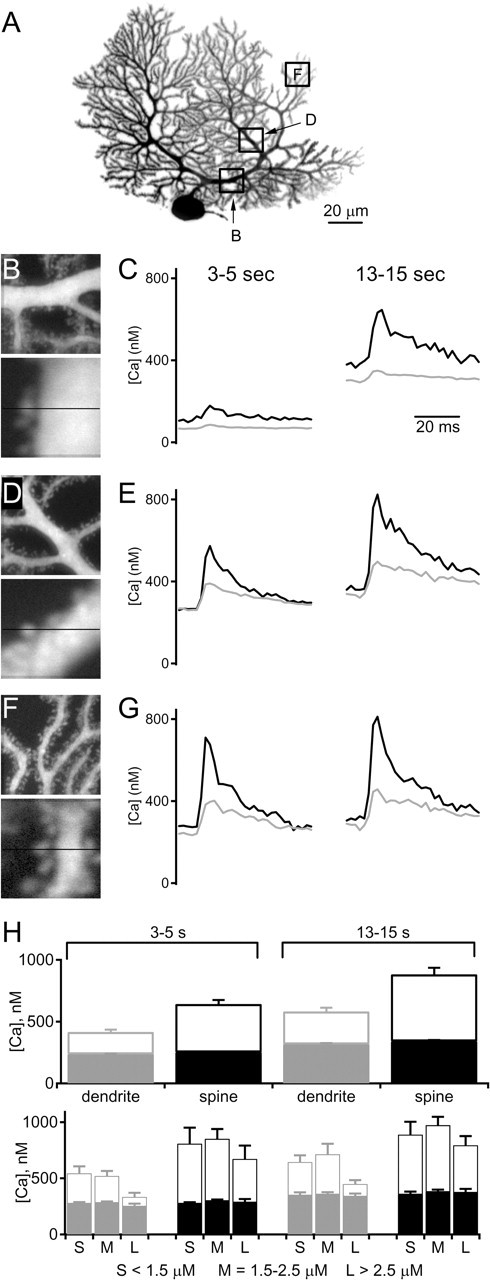
Endocannabinoids can act as retrograde messengers that allow postsynaptic cells to regulate the strength of their synaptic inputs. In the cerebellum, Purkinje cells (PCs) release endocannabinoids through two mechanisms. Synaptic activation evokes local endocannabinoid release that relies on a pathway that involves the metabotropic glutamate receptor mGluR1 and phospholipase-C (PLC). In contrast, depolarization evokes endocannabinoid release from the entire dendritic arbor. This leads to depolarization-induced suppression of inhibitory (DSI) and excitatory (DSE) synapses by a mechanism that does not involve mGluR1 or PLC. This latter mechanism of endocannabinoid release has only been observed under artificial conditions that transiently elevate postsynaptic calcium to >5 microm. Here, we tested the possibility that this mechanism could lead to retrograde inhibition in response to more realistic calcium signals. At both climbing fiber and inhibitory synapses onto PCs, we found that prolonging the elevation of calcium significantly lowered the peak calcium required to evoke PLC-independent endocannabinoid release. This suggests that the mechanism of endocannabinoid release involved in DSI and DSE is likely to evoke endocannabinoid release in response to physiologically relevant levels of calcium. When dendritic calcium was elevated to 0.4-1 microm for 15 s or more, endocannabinoid release from PCs selectively suppressed inhibitory synapses. This suggests that inhibitory synapses are more sensitive to prolonged calcium increases. Thus, in contrast to localized retrograde inhibition evoked by synaptic activation, modest but sustained calcium elevation could globally suppress inhibitory synapses onto PCs.
Figures
References
-
- Bacci A, Huguenard JR, Prince DA (2004). Long-lasting self-inhibition of neocortical interneurons mediated by endocannabinoids. Nature 431:312–316. - PubMed
-
- Brenowitz SD, Regehr WG (2005). Associative short-term synaptic plasticity mediated by endocannabinoids. Neuron 45:419–431. - PubMed
-
- Brown SP, Brenowitz SD, Regehr WG (2003). Brief presynaptic bursts evoke synapse-specific retrograde inhibition mediated by endogenous cannabinoids. Nat Neurosci 6:1048–1057. - PubMed
-
- Chevaleyre V, Castillo PE (2003). Heterosynaptic LTD of hippocampal GABAergic synapses: a novel role of endocannabinoids in regulating excitability. Neuron 38:461–472. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources