

Loading rat heart myocytes with Mg2+ using low-[Na+] solutions
- PMID: 16793904
- PMCID: PMC1819449
- DOI: 10.1113/jphysiol.2006.109850
Loading rat heart myocytes with Mg2+ using low-[Na+] solutions
Abstract
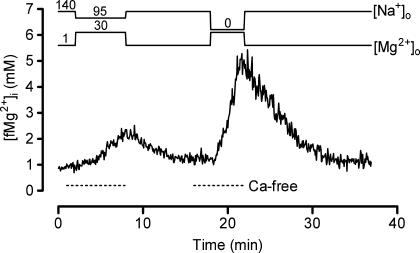
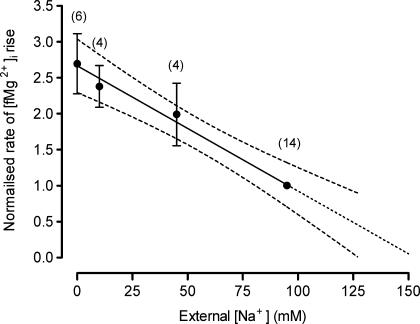
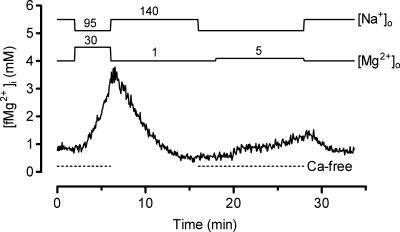
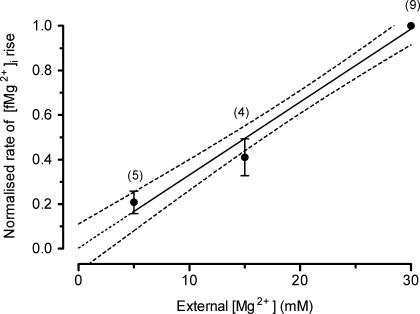
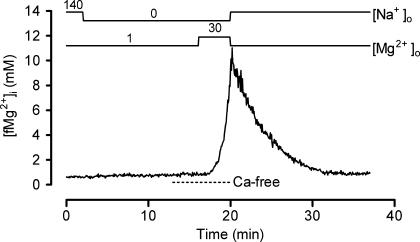
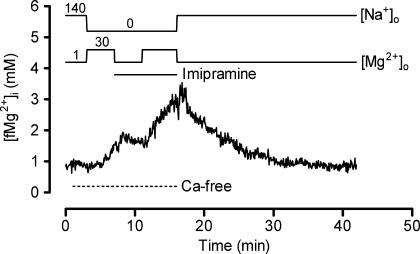
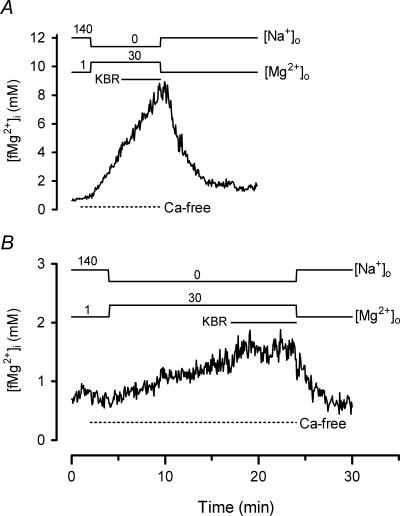
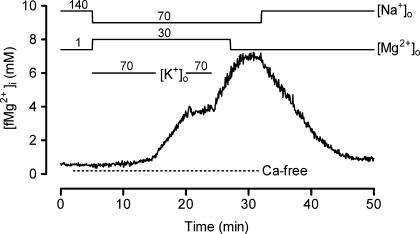
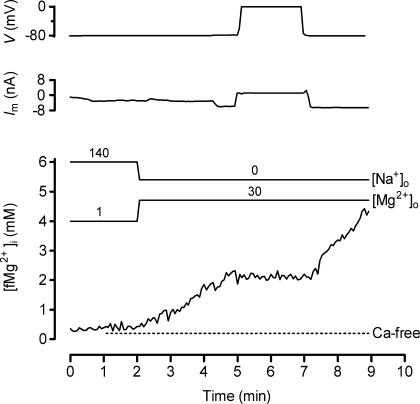
The objective of our study was to investigate how Mg2+ enters mammalian cardiac cells. During this work, we found evidence for a previously undescribed route for Mg2+ entry, and now provide a preliminary account of its properties. Changes in Mg2+ influx into rat ventricular myocytes were deduced from changes in intracellular ionized Mg2+ concentration ([fMg2+]i) measured from the fluorescence of mag-fura-2 loaded into isolated cells. Superfusion of myocytes at 37 degrees C with Ca2+-free solutions with both reduced [Na+] and raised [Mg2+] caused myocytes to load with Mg2+. Uptake was seen with solutions containing 5 mm Mg2+ and 95 mm Na+, and increased linearly with increasing extracellular [Mg2+] or decreasing extracellular [Na+]. It was very sensitive to temperature (Q(10) > 9, 25--37 degrees C), was observed even in myocytes with very low Na+ contents, and stopped abruptly when external [Na+] was returned to normal. Uptake was greatly reduced by imipramine or KB-R7943 if these were added when [fMg2+]i was close to the physiological level, but was unaffected if they were applied when [fMg2+]i was above 2 mm. Uptake was also reduced by depolarizing the membrane potential by increasing extracellular [K+] or voltage clamp to 0 mV. We suggest that initial Mg2+ uptake may involve several transporters, including reversed Na+-Mg2+ antiport and, depending on the exact conditions, reversed Na+-Ca2+ antiport. The ensuing rise of [fMg2+]i, in conjunction with reduced [Na+], may then activate a new Mg2+ transporter that is highly sensitive to temperature, is insensitive to imipramine or KB-R7943, but is inactivated by depolarization.
Figures
References
-
- Almulla HA, Bush PG, Steele MG, Flatman PW, Ellis D. Sodium-dependent recovery of ionised magnesium concentration following magnesium load in rat heart myocytes. Pflugers Arch. 2006;451:657–667. - PubMed
-
- Almulla HA, Ellis D, Flatman PW. Effects of imipramine and KB-R7943 on Na+ dependent Mg2+ transport in isolated rat ventricular myocytes. J Physiol. 2003;547.P:C19.
-
- Altura BM, Gebrewold A, Altura BT, Gupta RK. Role of brain [Mg2+]i in alcohol-induced hemorrhagic stroke in a rat model: a 31P-NMR in vivo study. Alcohol. 1995;12:131–136. - PubMed
-
- Buri A, Chen S, Fry CH, Illner H, Kickenweiz E, McGuigan JAS, Noble D, Powell T, Twist VW. The regulation of intracellular Mg2+ in guinea-pig heart, studied with Mg2+-selective microelectrodes and fluorochromes. Exp Physiol. 1993;78:221–233. - PubMed
-
- Cefaratti C, Romani A, Scarpa A. Differential localization and operation of distinct Mg2+ transporters in apical and basolateral sides of rat liver plasma membrane. J Biol Chem. 2000;275:3772–3780. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous