

Dopaminergic marker proteins in the substantia nigra of human immunodeficiency virus type 1-infected brains
- PMID: 16798675
- PMCID: PMC3710452
- DOI: 10.1080/13550280600724319
Dopaminergic marker proteins in the substantia nigra of human immunodeficiency virus type 1-infected brains
Abstract
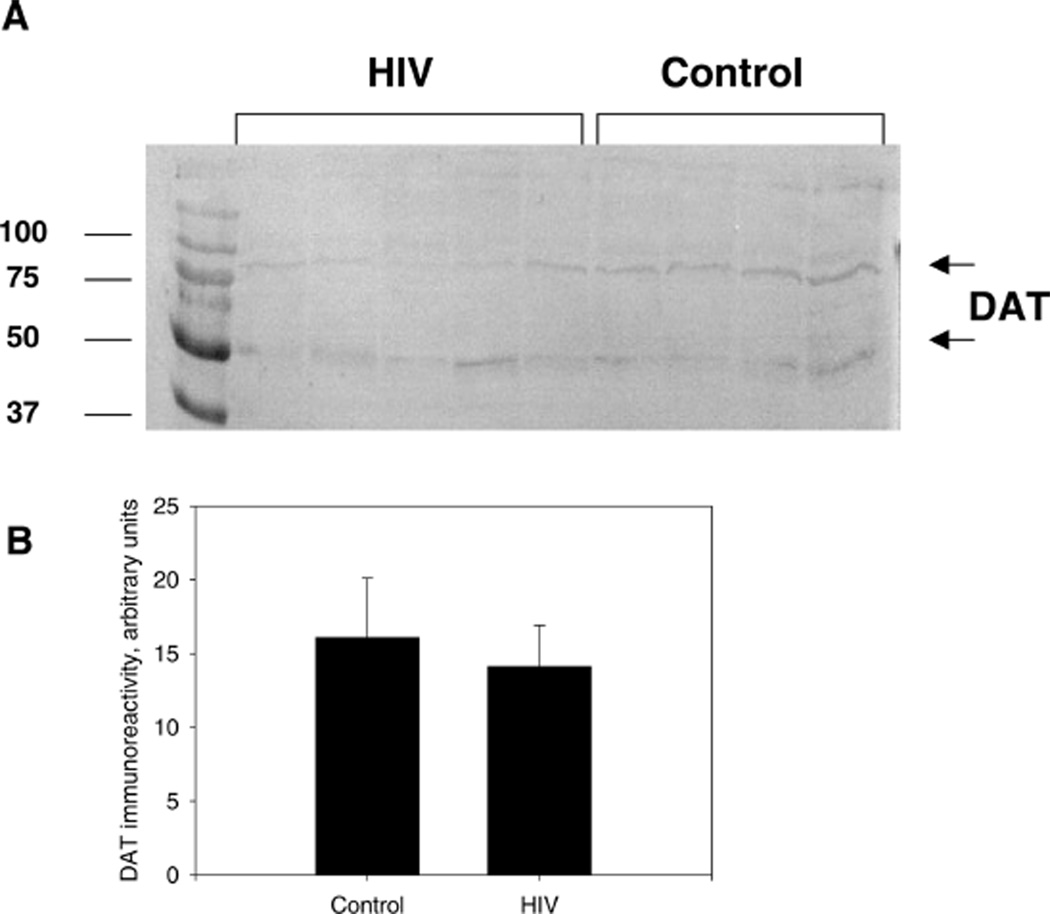
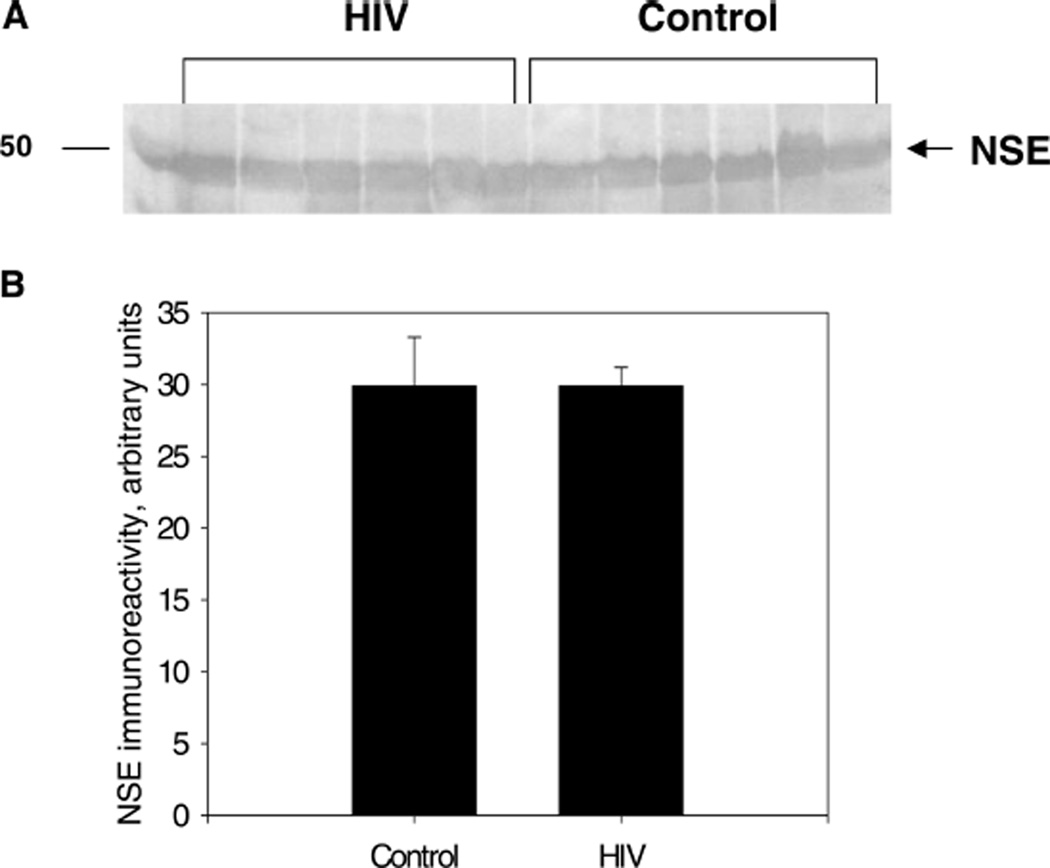
With the advent of highly active antiretroviral therapy, human immunodeficiency virus/acquired immunodeficiency syndrome (HIV/AIDS) is becoming a more chronic, manageable disease; nevertheless, the prevalence of neurological complications of AIDS is increasing. In this study, protein levels of tyrosine hydroxylase (TH) and dopamine transporter (DAT) in the substantia nigra of HIV-infected brains and -seronegative controls were determined by immunoblotting. The immunoreactivity of neuronal specific enolase (NSE) was used to assess cell loss. Although there were no changes in levels of immunoreactive DAT or NSE proteins in HIV brains, levels of immunoreactive TH were significantly reduced, relative to controls. These results suggest that decreases in TH, the rate-limiting enzyme of dopamine synthesis, may be a factor in the neurological manifestations of HIV infection.
Figures

References
-
- Aksenov MY, Hasselrot U, Wu G, Nath A, Anderson C, Mactutus CF, Booze RM. Temporal relationships between HIV-1 Tat-induced neuronal degeneration, OX-42 immunoreactivity, reactive astrocytosis, and protein oxidation in the rat striatum. Brain Res. 2003;987:1–9. - PubMed
-
- Arendt G. Neurological manifestations of HIV-infection in the era of highly active antiretroviral therapy (HAART) Fortschr Neurol Psychiatr. 2005;73:577–586. - PubMed
-
- Berger JR, Arendt G. HIV dementia: the role of the basal ganglia and dopaminergic systems. Psychopharmacol. 2000;14:214–221. - PubMed
-
- Blanchard-Fillion B, Souza JM, Friel T, Jiang GC, Vrana K, Sharov V, et al. Nitration and inactivation of tyrosine hydroxylase by peroxynitrite. J Biol Chem. 2001;279:46017–46023. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
