

A burst of auxilin recruitment determines the onset of clathrin-coated vesicle uncoating
- PMID: 16798879
- PMCID: PMC1502446
- DOI: 10.1073/pnas.0603369103
A burst of auxilin recruitment determines the onset of clathrin-coated vesicle uncoating
Abstract
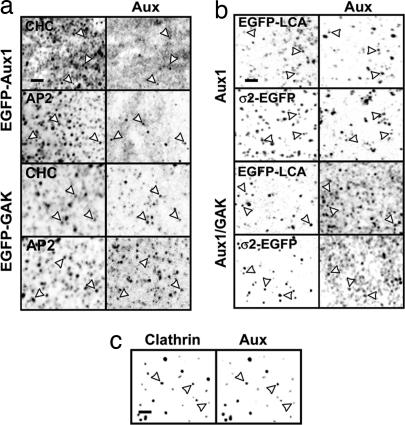
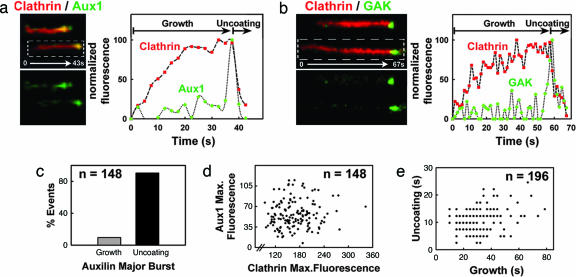
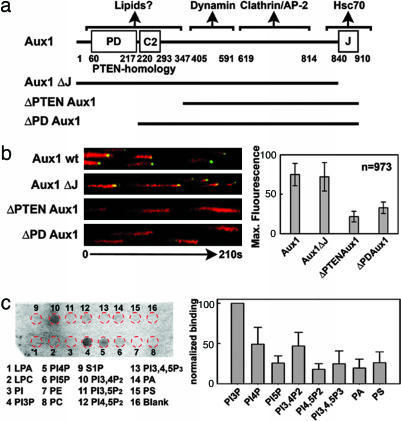
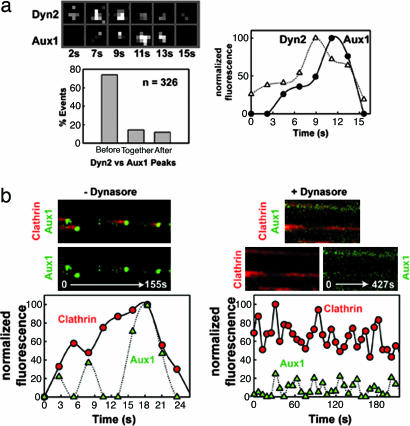
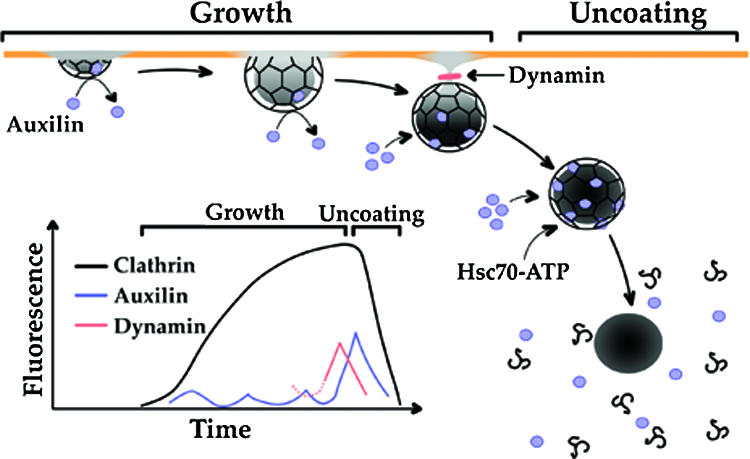
Clathrin-coated pits assemble on a membrane and pinch off as coated vesicles. The released vesicles then rapidly lose their clathrin coats in a process mediated by the ATPase Hsc70, recruited by auxilin, a J-domain-containing cofactor. How is the uncoating process regulated? We find that during coat assembly small and variable amounts of auxilin are recruited transiently but that a much larger burst of association occurs after the peak of dynamin signal, during the transition between membrane constriction and vesicle budding. We show that the auxilin burst depends on domains of the protein likely to interact with lipid head groups. We conclude that the timing of auxilin recruitment determines the onset of uncoating. We propose that, when a diffusion barrier is established at the constricting neck of a fully formed coated pit and immediately after vesicle budding, accumulation of a specific lipid can recruit sufficient auxilin molecules to trigger uncoating.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
-
- Ehrlich M., Boll W., Van Oijen A., Hariharan R., Chandran K., Nibert M. L., Kirchhausen T. Cell. 2004;118:591–605. - PubMed
-
- Lemmon S. K. Curr. Biol. 2001;11:R49–R52. - PubMed
-
- Greener T., Zhao X., Nojima H., Eisenberg E., Greene L. E. J. Biol. Chem. 2000;275:1365–1370. - PubMed
-
- Jiang J. W., Taylor A. B., Prasad K., Ishikawa-Brush Y., Hart P. J., Lafer E. M., Sousa R. Biochemistry. 2003;42:5748–5753. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
