

Deletion of the neuropeptide Y (NPY) Y1 receptor gene reveals a regulatory role of NPY on catecholamine synthesis and secretion
- PMID: 16798884
- PMCID: PMC1502486
- DOI: 10.1073/pnas.0600913103
Deletion of the neuropeptide Y (NPY) Y1 receptor gene reveals a regulatory role of NPY on catecholamine synthesis and secretion
Abstract
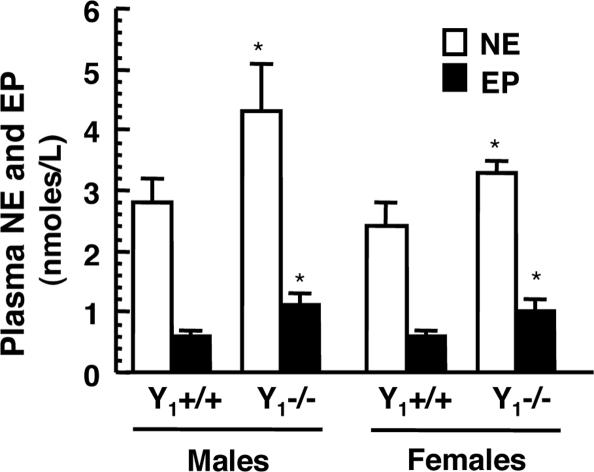
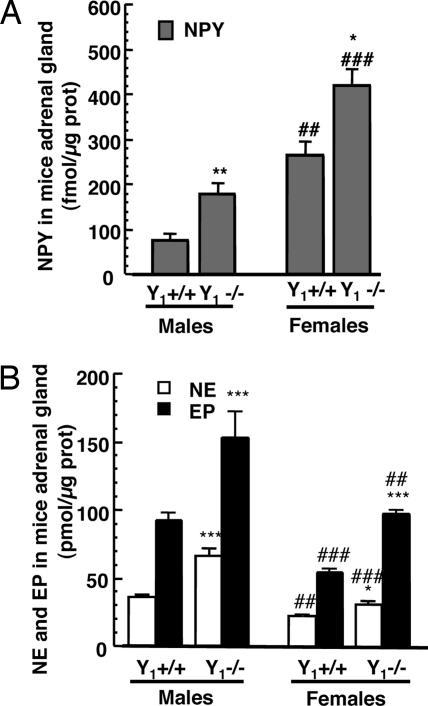
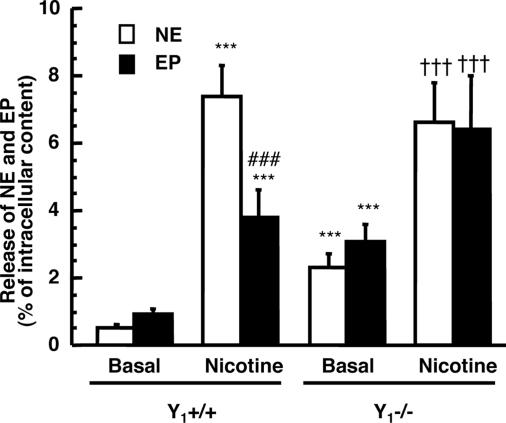
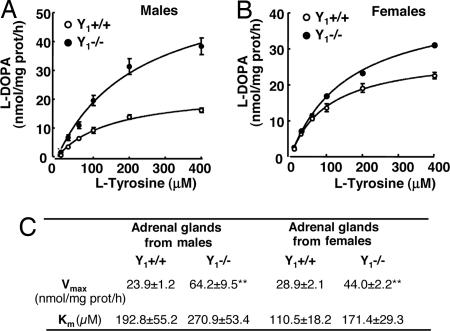
The contribution of neuropeptide Y (NPY), deriving from adrenal medulla, to the adrenosympathetic tone is unknown. We found that in response to NPY, primary cultures of mouse adrenal chromaffin cells secreted catecholamine, and that this effect was abolished in cultures from NPY Y(1) receptor knockout mice (Y(1)-/-). Compared with wild-type mice (Y(1)+/+), the adrenal content and constitutive release of catecholamine were increased in chromaffin cells from Y(1)-/- mice. In resting animals, catecholamine plasma concentrations were higher in Y(1)-/- mice. Comparing the adrenal glands of both genotypes, no differences were observed in the area of the medulla, cortex, and X zone. The high turnover of adrenal catecholamine in Y(1)-/- mice was explained by the enhancement of tyrosine hydroxylase (TH) activity, although no change in the affinity of the enzyme was observed. The molecular interaction between the Y(1) receptor and TH was demonstrated by the fact that NPY markedly inhibited the forskolin-induced luciferin activity in Y(1) receptor-expressing SK-N-MC cells transfected with a TH promoter sequence. We propose that NPY controls the release and synthesis of catecholamine from the adrenal medulla and consequently contributes to the sympathoadrenal tone.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures




References
-
- Lundberg J. M., Torssell L., Sollevi A., Pernow J., Theodorsson Norheim E., Anggard A., Hamberger B. Regul. Pept. 1985;13:41–52. - PubMed
-
- Takiyyuddin M. A., Brown M. R., Dinh T. Q., Cervenka J. H., Braun S. D., Parmer R. J., Kennedy B., O’Connor D. T. J. Auton. Pharmacol. 1994;14:187–200. - PubMed
-
- Michel M. C., Beck-Sickinger A., Cox H., Doods H. N., Herzog H., Larhammar D., Quirion R., Schwartz T., Westfall T. Pharmacol. Rev. 1998;50:143–150. - PubMed
-
- Silva A. P., Cavadas C., Grouzmann E. Clin. Chim. Acta. 2002;326:3–25. - PubMed
-
- Walker P., Grouzmann E., Burnier M., Waeber B. Trends Pharmacol. Sci. 1991;12:111–115. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
