

Endocytic vesicle scission by lipid phase boundary forces
- PMID: 16801551
- PMCID: PMC1502448
- DOI: 10.1073/pnas.0601045103
Endocytic vesicle scission by lipid phase boundary forces
Abstract
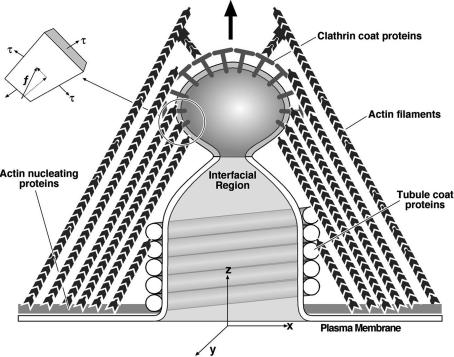
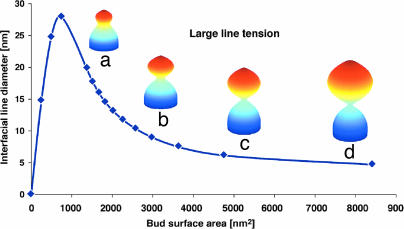
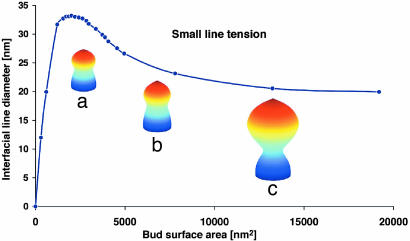
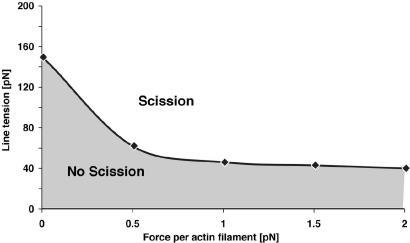
Endocytosis in budding yeast is thought to occur in several phases. First, the membrane invaginates and then elongates into a tube. A vesicle forms at the end of the tube, eventually pinching off to form a "free" vesicle. Experiments show that actin polymerization is an active participant in the endocytic process, along with a number of membrane-associated proteins. Here we investigate the possible roles of these components in driving vesiculation by constructing a quantitative model of the process beginning at the stage where the membrane invagination has elongated into a tube encased in a sheath of membrane-associated protein. This protein sheath brings about the scission step where the vesicle separates from the tube. When the protein sheath is dynamin, it is commonly assumed that scission is brought about by the constriction of the sheath. Here, we show that an alternative scenario can work as well: The protein sheath acts as a "filter" to effect a phase separation of lipid species. The resulting line tension tends to minimize the interface between the tube region and the vesicle region. Interestingly, large vesicle size can further facilitate the reduction of the interfacial diameter down to a few nanometers, small enough so that thermal fluctuations can fuse the membrane and pinch off the vesicle. To deform the membrane into the tubular vesicle shape, the membrane elastic resistance forces must be balanced by some additional forces that we show can be generated by actin polymerization and/or myosin I. These active forces are shown to be important in successful scission processes as well.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
Similar articles
-
Direct and remote constriction of membrane necks.Phys Rev E Stat Nonlin Soft Matter Phys. 2014 May;89(5):052701. doi: 10.1103/PhysRevE.89.052701. Epub 2014 May 2. Phys Rev E Stat Nonlin Soft Matter Phys. 2014. PMID: 25353823
-
A dynamin-actin interaction is required for vesicle scission during endocytosis in yeast.Curr Biol. 2015 Mar 30;25(7):868-78. doi: 10.1016/j.cub.2015.01.061. Epub 2015 Mar 12. Curr Biol. 2015. PMID: 25772449 Free PMC article.
-
Actin and endocytosis in budding yeast.Genetics. 2015 Feb;199(2):315-58. doi: 10.1534/genetics.112.145540. Genetics. 2015. PMID: 25657349 Free PMC article. Review.
-
Mechanochemical crosstalk during endocytic vesicle formation.Curr Opin Cell Biol. 2010 Feb;22(1):36-43. doi: 10.1016/j.ceb.2009.11.009. Epub 2009 Dec 21. Curr Opin Cell Biol. 2010. PMID: 20022735 Free PMC article.
-
SNX9 - a prelude to vesicle release.J Cell Sci. 2009 Jan 1;122(Pt 1):5-11. doi: 10.1242/jcs.037135. J Cell Sci. 2009. PMID: 19092055 Review.
Cited by
-
Cardiolipin's propensity for phase transition and its reorganization by dynamin-related protein 1 form a basis for mitochondrial membrane fission.Mol Biol Cell. 2015 Sep 1;26(17):3104-16. doi: 10.1091/mbc.E15-06-0330. Epub 2015 Jul 8. Mol Biol Cell. 2015. PMID: 26157169 Free PMC article.
-
Rab and actomyosin-dependent fission of transport vesicles at the Golgi complex.Nat Cell Biol. 2010 Jul;12(7):645-54. doi: 10.1038/ncb2067. Epub 2010 Jun 20. Nat Cell Biol. 2010. PMID: 20562865
-
PtdIns(4,5)P2 turnover is required for multiple stages during clathrin- and actin-dependent endocytic internalization.J Cell Biol. 2007 Apr 23;177(2):355-67. doi: 10.1083/jcb.200611011. J Cell Biol. 2007. PMID: 17452534 Free PMC article.
-
Ras Multimers on the Membrane: Many Ways for a Heart-to-Heart Conversation.Genes (Basel). 2022 Jan 25;13(2):219. doi: 10.3390/genes13020219. Genes (Basel). 2022. PMID: 35205266 Free PMC article. Review.
-
Clathrin-independent pathways of endocytosis.Cold Spring Harb Perspect Biol. 2014 Jun 2;6(6):a016758. doi: 10.1101/cshperspect.a016758. Cold Spring Harb Perspect Biol. 2014. PMID: 24890511 Free PMC article.
References
-
- Conner S. D., Schmid S. L. Nature. 2003;422:37–44. - PubMed
-
- Higgins M. K., McMahon H. T. Trends Biochem. Sci. 2002;27:257–263. - PubMed
-
- Merrifield C. J. Trends Cell Biol. 2004;14:352–358. - PubMed
-
- Praefcke G. J., McMahon H. T. Nat. Rev. Mol. Cell Biol. 2004;5:133–147. - PubMed
-
- Song B. D., Schmid S. L. Biochemistry. 2003;42:1369–1376. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
