

Regulation of poly(A) binding protein function in translation: Characterization of the Paip2 homolog, Paip2B
- PMID: 16804161
- PMCID: PMC1524897
- DOI: 10.1261/rna.106506
Regulation of poly(A) binding protein function in translation: Characterization of the Paip2 homolog, Paip2B
Abstract
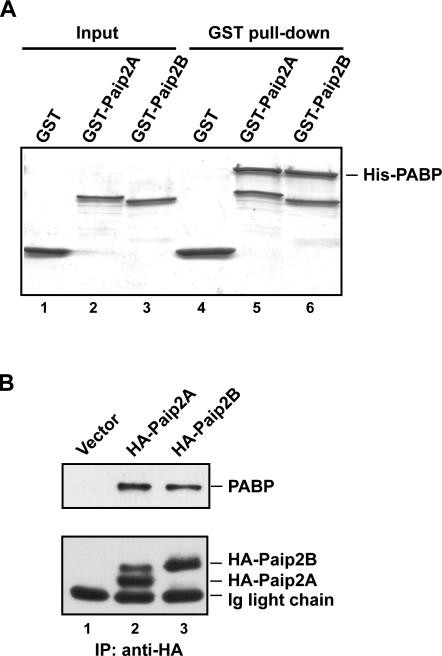
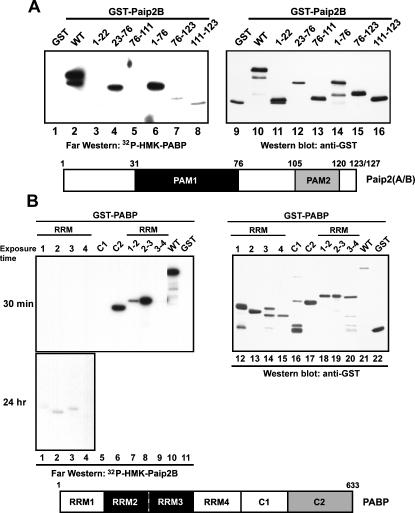
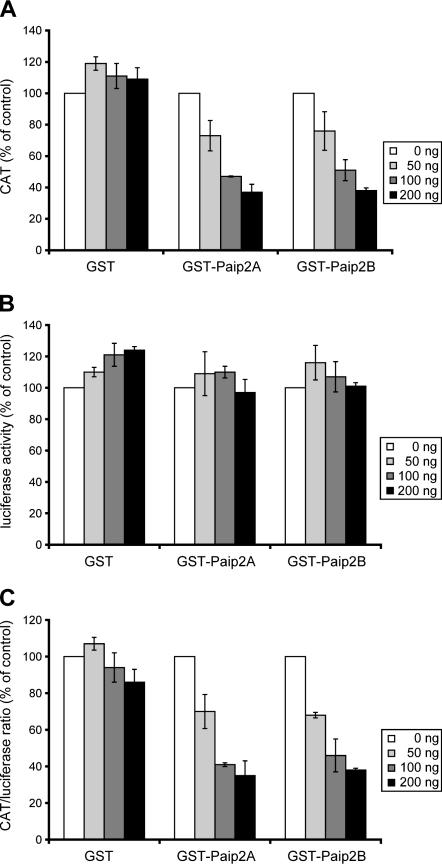
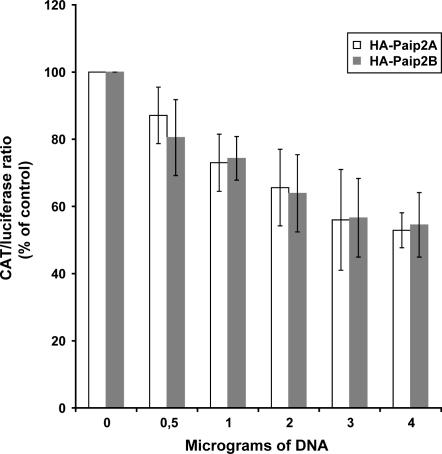
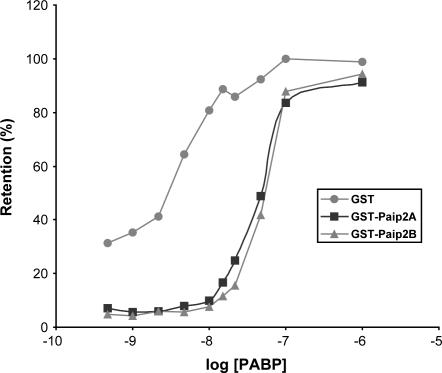
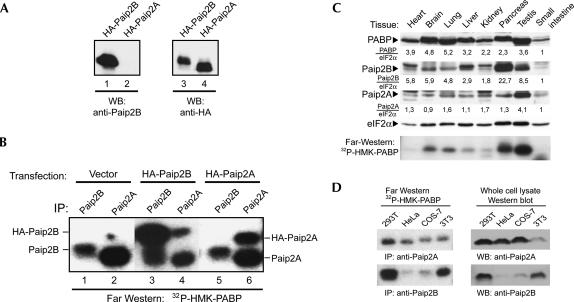
The 5' cap and 3' poly(A) tail of eukaryotic mRNAs act synergistically to enhance translation. This synergy is mediated via interactions between eIF4G (a component of the eIF4F cap binding complex) and poly(A) binding protein (PABP). Paip2 (PABP-interacting protein 2) binds PABP and inhibits translation both in vitro and in vivo by decreasing the affinity of PABP for polyadenylated RNA. Here, we describe the functional characteristics of Paip2B, a Paip2 homolog. A full-length brain cDNA of Paip2B encodes a protein that shares 59% identity and 80% similarity with Paip2 (Paip2A), with the highest conservation in the two PABP binding domains. Paip2B acts in a manner similar to Paip2A to inhibit translation of capped and polyadenylated mRNAs both in vitro and in vivo by displacing PABP from the poly(A) tail. Also, similar to Paip2A, Paip2B does not affect the translation mediated by the internal ribosome entry site (IRES) of hepatitis C virus (HCV). However, Paip2A and Paip2B differ with respect to both mRNA and protein distribution in different tissues and cell lines. Paip2A is more highly ubiquitinated than is Paip2B and is degraded more rapidly by the proteasome. Paip2 protein degradation may constitute a primary mechanism by which cells regulate PABP activity in translation.
Figures



References
-
- Altschul S.F., Gish W., Miller W., Myers E.W., Lipman D.J. Basic local alignment search tool. J. Mol. Biol. 1990;215:403–410. - PubMed
-
- Blanar M.A., Rutter W.J. Interaction cloning: Identification of a helix-loop-helix zipper protein that interacts with c-Fos. Science. 1992;256:1014–1018. - PubMed
-
- Bravo J., Aguilar-Henonin L., Olmedo G., Guzman P. Four distinct classes of proteins as interaction partners of the PABC domain of Arabidopsis thaliana Poly(A)-binding proteins. Mol. Genet. Genomics. 2005;272:651–665. - PubMed
-
- Callaghan M.J., Russell A.J., Woollatt E., Sutherland G.R., Sutherland R.L., Watts C.K. Identification of a human HECT family protein with homology to the Drosophila tumor suppressor gene hyperplastic discs. Oncogene. 1998;17:3479–3491. - PubMed
-
- Combet C., Blanchet C., Geourjon C., Deleage G. NPS@: Network protein sequence analysis. Trends Biochem. Sci. 2000;25:147–150. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous