

Downregulation of uPA, uPAR and MMP-9 using small, interfering, hairpin RNA (siRNA) inhibits glioma cell invasion, angiogenesis and tumor growth
- PMID: 16804563
- PMCID: PMC1483066
- DOI: 10.1017/s1740925x04000237
Downregulation of uPA, uPAR and MMP-9 using small, interfering, hairpin RNA (siRNA) inhibits glioma cell invasion, angiogenesis and tumor growth
Abstract
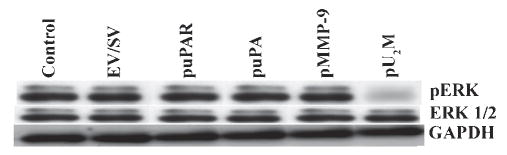
The diffuse, extensive infiltration of malignant gliomas into the surrounding normal brain is believed to rely on modification of the proteolysis of extracellular matrix components. Our previous results clearly demonstrate that uPA, uPAR and MMP-9 concentrations increase significantly during tumor progression and that tumor growth can be inhibited with antisense stable clones of these molecules. Because antisense-mediated gene silencing does not completely inhibit the translation of target mRNA and high concentrations of antisense molecules are required to achieve gene silencing, we used the RNAi approach to silence uPA, uPAR and MMP-9 in this study. We examined a cytomegalovirus (CMV) promoter-driven DNA-template approach to induce hairpin RNA (hpRNA)-triggered RNAi to inhibit uPA, uPAR and MMP-9 gene expression with a single construct. uPAR protein levels and enzymatic activity of uPA and MMP-9 were found to significantly decrease in cells transfected with a plasmid expressing hairpin siRNA for uPAR, uPA and MMP-9. pU(2)M-transfected SNB19 cells significantly decreased uPA, uPAR and MMP-9 expression compared to mock and EV/SV-transfected cells, determined by immunohistochemical analysis. Furthermore, the effect of the single constructs for these molecules was a specific inhibition of their respective protein levels, as demonstrated by immunohistochemical analysis. After transfection with a plasmid vector expressing dsRNA for uPA, uPAR and MMP-9, glioma-cell invasion was retarded compared with mock and EV/SV-treated groups, demonstrated by Matrigel-invasion assay and spheroid-invasion assay. Downregulation of uPA, uPAR and MMP-9 using RNAi inhibited angiogenesis in an in vitro (co-culture) model. Direct intratumoral injections of plasmid DNA expressing hpRNA for uPA, uPAR and MMP-9 significantly regressed pre-established intracranial tumors in nude mice. In addition, cells treated with RNAi for uPAR, uPA and MMP-9 showed reduced pERK levels compared with parental and EV/SV-treated SNB19 cells. Our results support the therapeutic potential of RNAi as a method for gene therapy in treating gliomas.
Figures
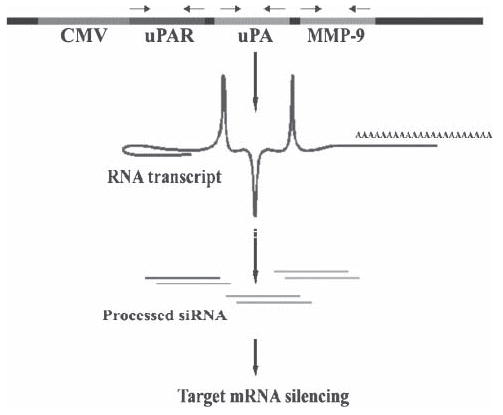
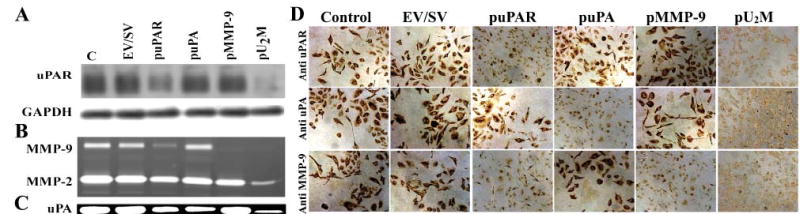
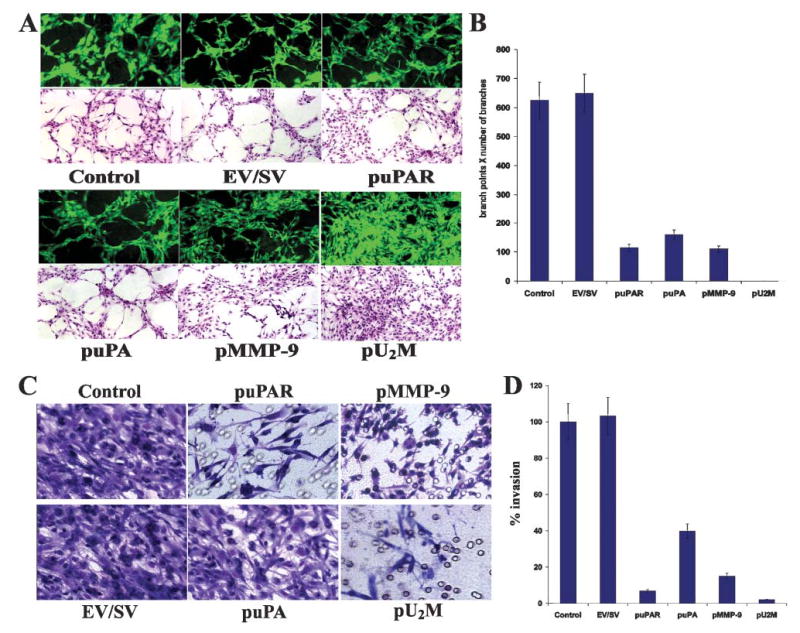
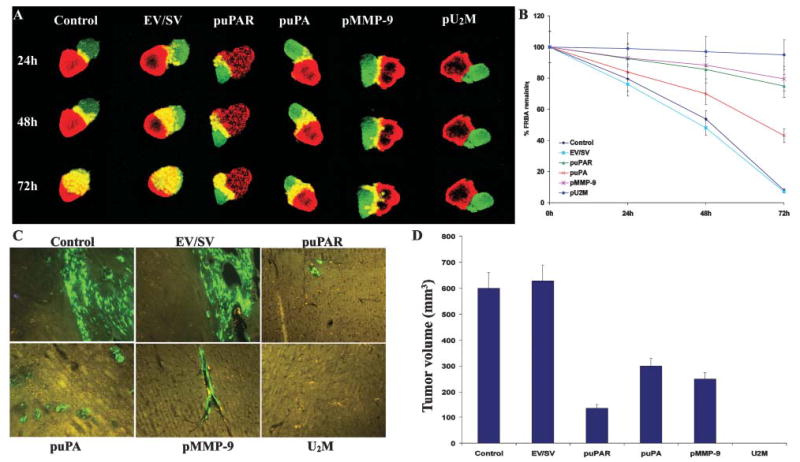
References
-
- Alemany R, Gomez-Manzano C, Balague C, Yung WK, Curiel DT, Kyritsis AP, Fueyo J. Gene therapy for gliomas: molecular targets, adenoviral vectors, and oncolytic adenoviruses. Experimental Cell Research. 1999;252:1–12. - PubMed
-
- Alonso DF, Farias EF, Ladeda V, Davel L, Puricelli L, Bal de Kier JE. Effects of synthetic urokinase inhibitors on local invasion and metastasis in a murine mammary tumor model. Breast Cancer Research Treatment. 1996;40:209–223. - PubMed
-
- Alonso DF, Tejera AM, Farias EF, Bal de Kier JE, Gomez DE. Inhibition of mammary tumor cell adhesion, migration, and invasion by the selective synthetic urokinase inhibitor B428. Anticancer Research. 1998;18:4499–4504. - PubMed
-
- Andreasen PA, Kjoller L, Christensen L, Duffy MJ. The urokinase-type plasminogen activator system in cancer metastasis: a review. International Journal of Cancer. 1997;72:1–22. - PubMed
-
- Behzadian MA, Windsor LJ, Ghaly N, Liou G, Tsai NT, Caldwell RB. VEGF-induced paracellular permeability in cultured endothelial cells involves urokinase and its receptor. FASEB Journal. 2003;17:752–754. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous