

N-cadherin transsynaptically regulates short-term plasticity at glutamatergic synapses in embryonic stem cell-derived neurons
- PMID: 16807326
- PMCID: PMC6673917
- DOI: 10.1523/JNEUROSCI.1013-06.2006
N-cadherin transsynaptically regulates short-term plasticity at glutamatergic synapses in embryonic stem cell-derived neurons
Abstract
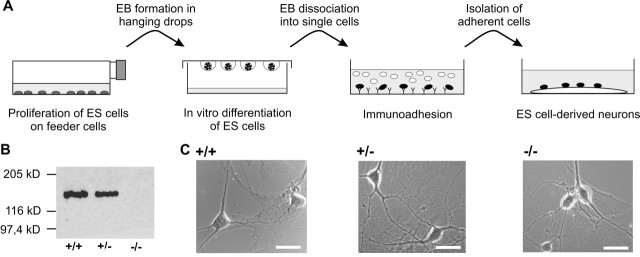
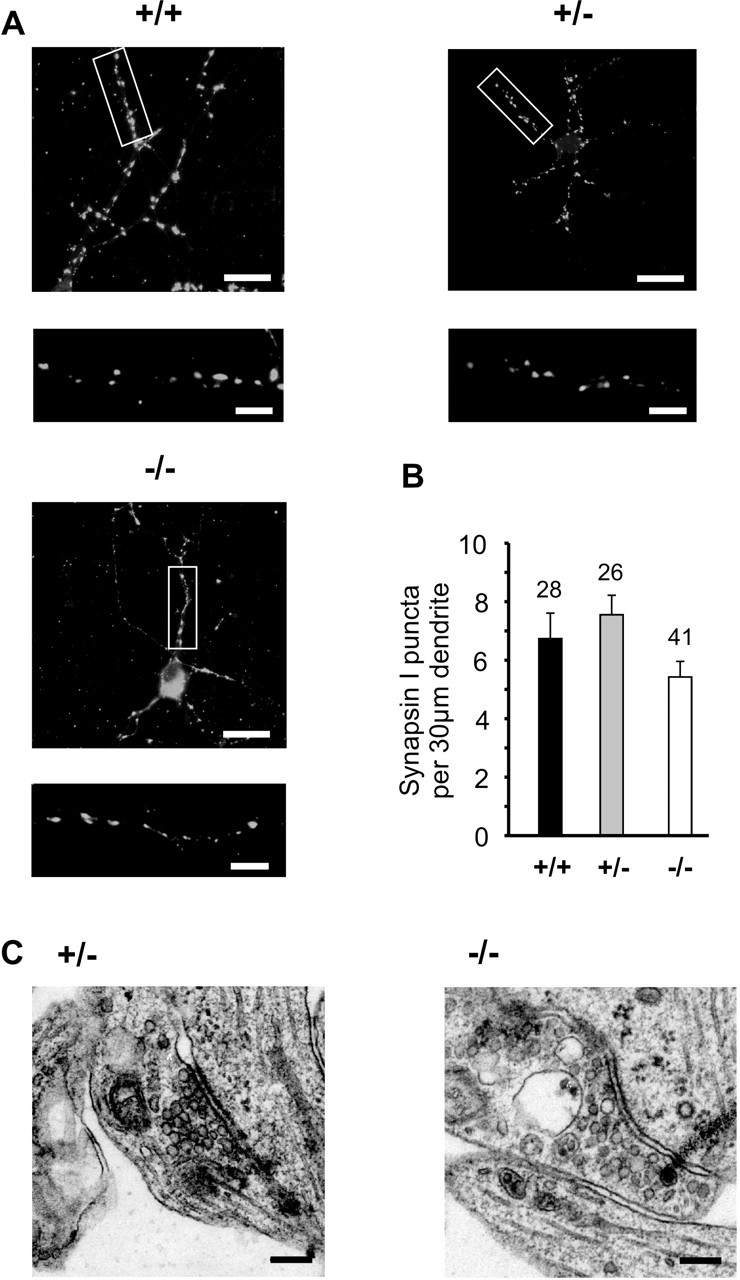
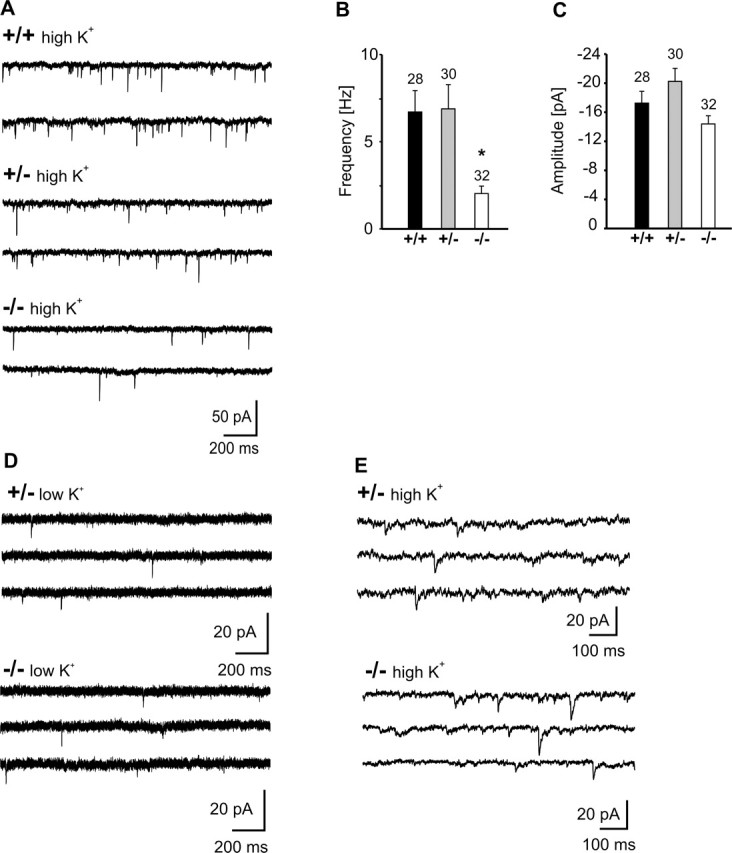
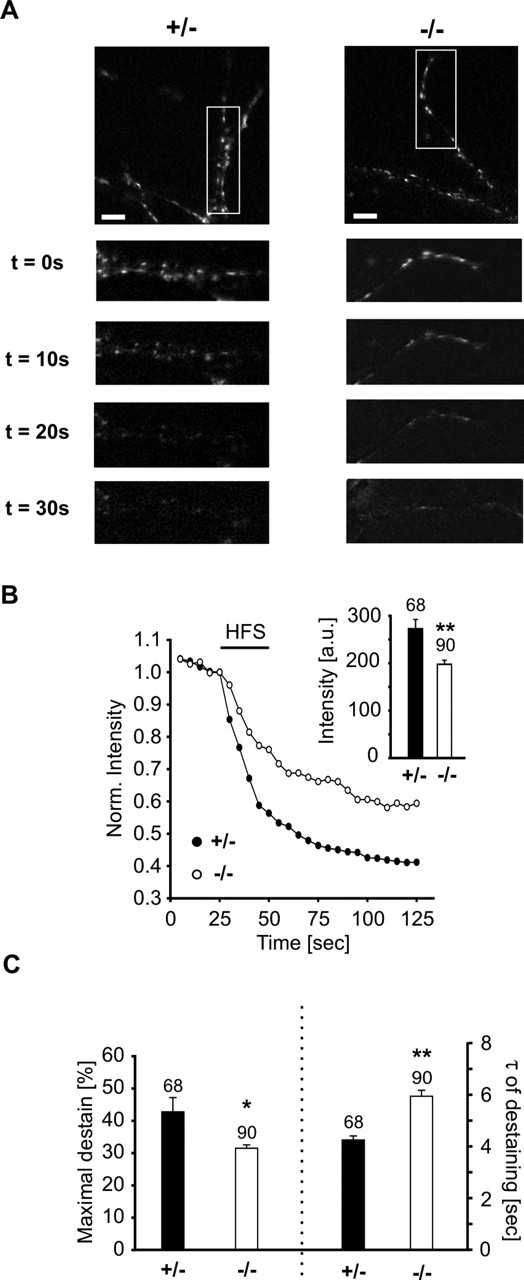
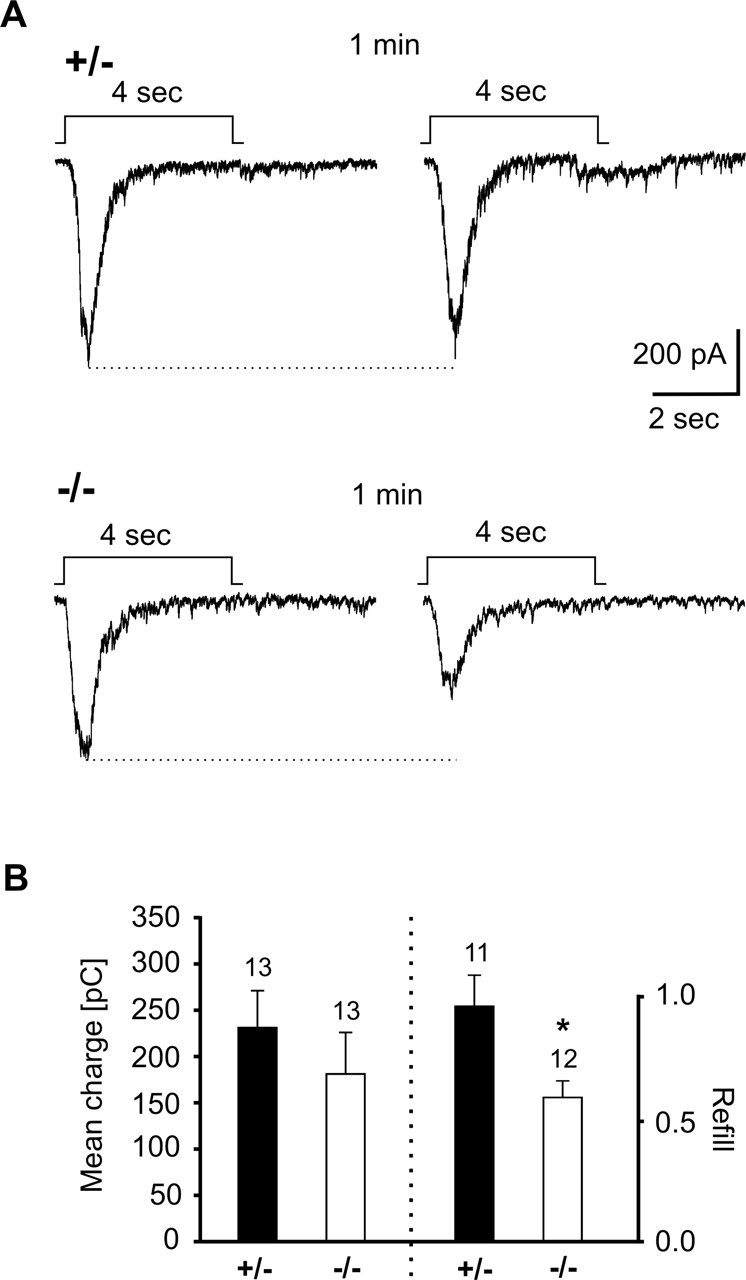
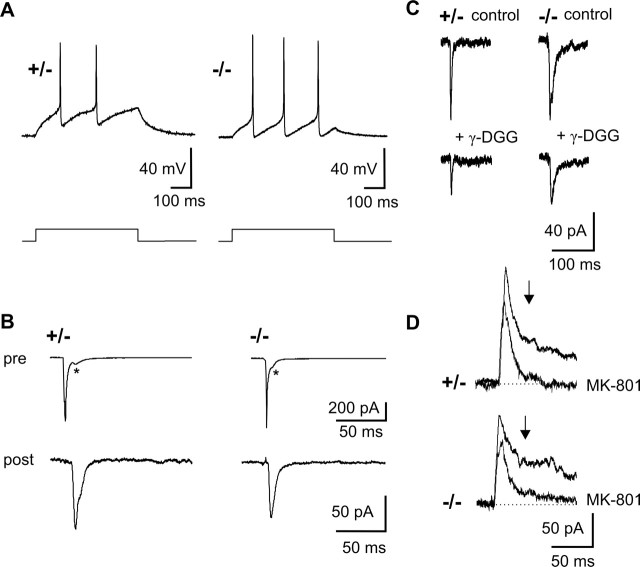
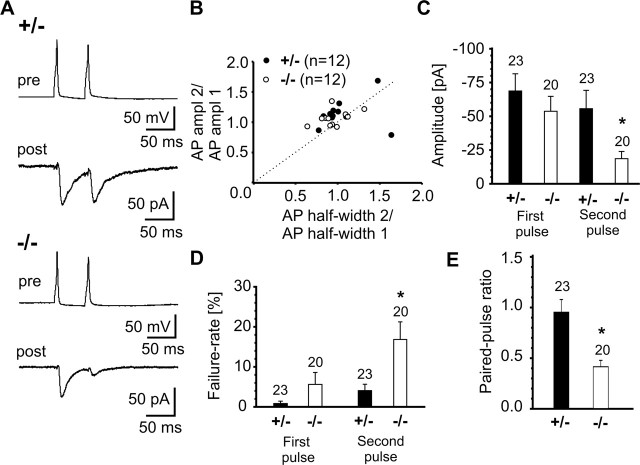
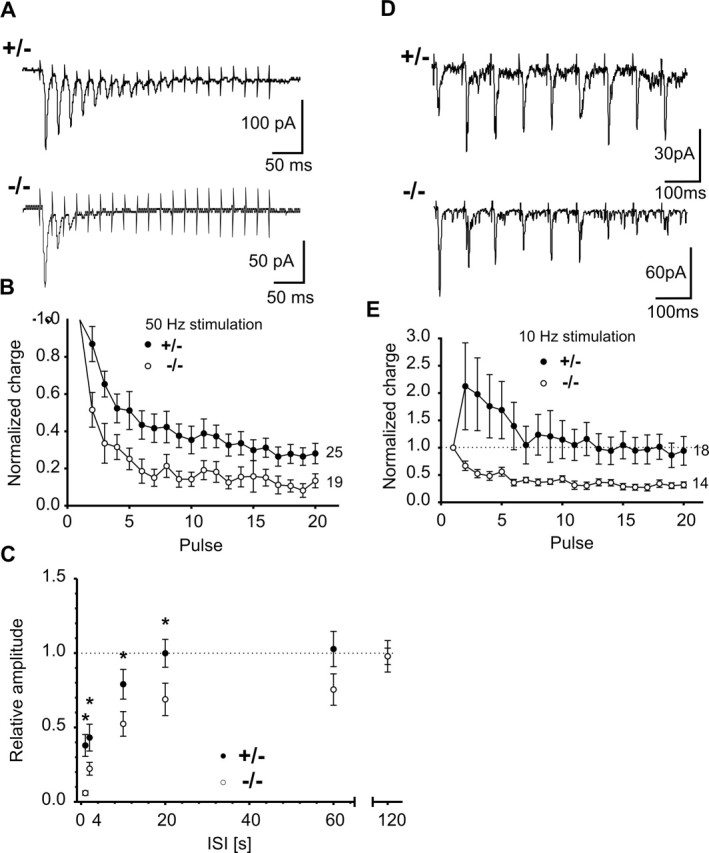
The cell adhesion molecule N-cadherin has been proposed to regulate synapse formation in mammalian central neurons. This is based on its synaptic localization enabling alignment of presynaptic and postsynaptic specializations by an adhesion mechanism. However, a potential role of N-cadherin in regulating synaptic transmission has remained elusive. In this paper, a functional analysis of N-cadherin knock-out synapses was enabled by in vitro neuronal differentiation of mouse embryonic stem cells circumventing the early embryonic lethality of mice genetically null for N-cadherin. In our in vitro system, initial synapse formation was not altered in the absence of N-cadherin, which might be attributable to compensatory mechanisms. Here, we demonstrate that N-cadherin is required for regulating presynaptic function at glutamatergic synapses. An impairment in the availability of vesicles for exocytosis became apparent selectively during high activity. Short-term plasticity was strongly altered with synaptic depression enhanced in the absence of N-cadherin. Most intriguingly, facilitation was converted to depression under specific stimulation conditions. This indicates an important role of N-cadherin in the control of short-term plasticity. To analyze, whether N-cadherin regulates presynaptic function by a transsynaptic mechanism, we studied chimeric cultures consisting of wild-type neocortical neurons and ES cell-derived neurons. With N-cadherin absent only postsynaptically, we observed a similar increase in short-term synaptic depression as found in its complete absence. This indicates a retrograde control of short-term plasticity by N-cadherin. In summary, our results revealed an unexpected involvement of a synaptic adhesion molecule in the regulation of short-term plasticity at glutamatergic synapses.
Figures
References
-
- Abe K, Chisaka O, van Roy F, Takeichi M (2004). Stability of dendritic spines and synaptic contacts is controlled by αN-catenin. Nat Neurosci 7:357–363. - PubMed
-
- Angst BD, Marcozzi C, Magee AI (2001). The cadherin superfamily: diversity in form and function. J Cell Sci 114:629–641. - PubMed
-
- Bozdagi O, Shan W, Tanaka H, Benson DL, Huntley GW (2000). Increasing numbers of synaptic puncta during late-phase LTP: N-cadherin is synthesized, recruited to synaptic sites, and required for potentiation. Neuron 28:245–259. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials