

Preferential infection shortens the life span of human immunodeficiency virus-specific CD4+ T cells in vivo
- PMID: 16809286
- PMCID: PMC1489023
- DOI: 10.1128/JVI.00070-06
Preferential infection shortens the life span of human immunodeficiency virus-specific CD4+ T cells in vivo
Abstract
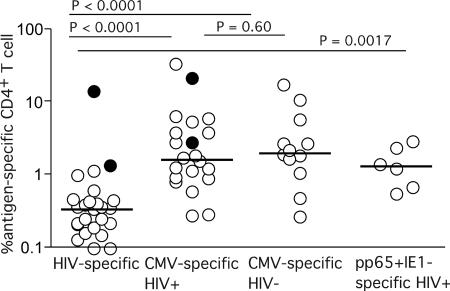
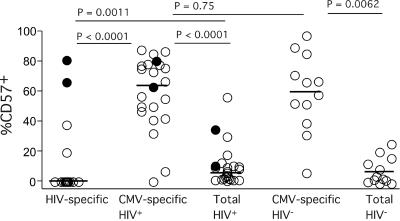
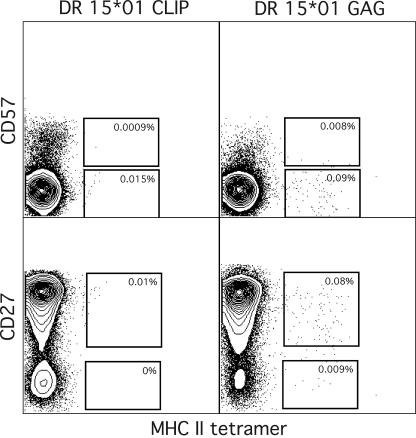
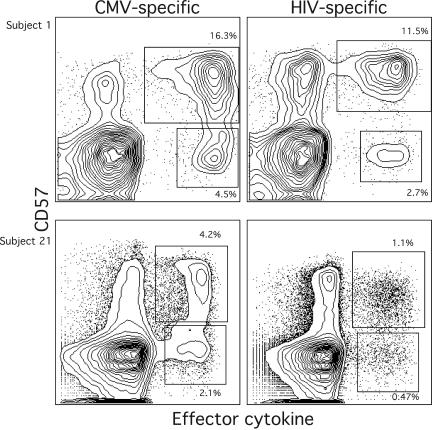
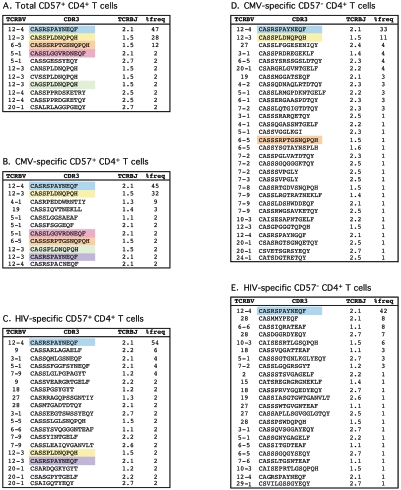
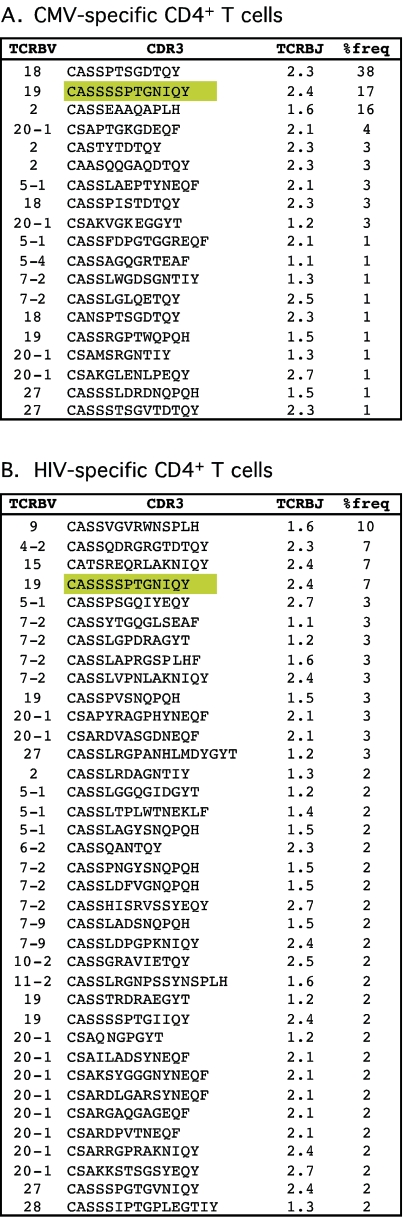
CD4(+) T-cell help is essential for effective immune responses to viruses. In human immunodeficiency virus (HIV) infection, CD4(+) T cells specific for HIV are infected by the virus at higher frequencies than other memory CD4(+) T cells. Here, we demonstrate that HIV-specific CD4(+) T cells are barely detectable in most infected individuals and that the corresponding CD4(+) T cells exhibit an immature phenotype compared to both cytomegalovirus (CMV)-specific CD4(+) T cells and other memory CD4(+) T cells. However, in two individuals, we observed a rare and diametrically opposed pattern in which HIV-specific CD4(+) T-cell populations of large magnitude exhibited a terminally differentiated immunophenotype; these cells were not preferentially infected in vivo. Clonotypic analysis revealed that the HIV-specific CD4(+) T cells from these individuals were cross-reactive with CMV. Thus, preferential infection can be circumvented in the presence of cross-reactive CD4(+) T cells driven to maturity by coinfecting viral antigens, and this physical proximity rather than activation status per se is an important determinant of preferential infection based on antigen specificity. These data demonstrate that preferential infection reduces the life span of HIV-specific CD4(+) T cells in vivo and thereby compromises the generation of effective immune responses to the virus itself; further, this central feature in the pathophysiology of HIV infection can be influenced by the cross-reactivity of responding CD4(+) T cells.
Figures
References
-
- Betts, M. R., D. R. Ambrozak, D. C. Douek, S. Bonhoeffer, J. M. Brenchley, J. P. Casazza, R. A. Koup, and L. J. Picker. 2001. Analysis of total human immunodeficiency virus (HIV)-specific CD4+ and CD8+ T-cell responses: relationship to viral load in untreated HIV infection. J. Virol. 75:11983-11991. - PMC - PubMed
-
- Betts, M. R., J. P. Casazza, B. A. Patterson, S. Waldrop, W. Trigona, T.-M. Fu, F. Kern, L. J. Picker, and R. A. Koup. 2000. Putative immunodominant human immunodeficiency virus-specific CD8+ T-cell responses cannot be predicted by major histocompatibility complex class I haplotype. J. Virol. 74:9144-9151. - PMC - PubMed
-
- Birch, M. R., J. C. Learmont, W. B. Dyer, N. J. Deacon, J. J. Zaunders, N. Saksena, A. L. Cunningham, J. Mills, and J. S. Sullivan. 2001. An examination of signs of disease progression in survivors of the Sydney Blood Bank Cohort (SBBC). J. Clin. Virol. 22:263-270. - PubMed
-
- Boaz, M. J., A. Waters, S. Murad, P. J. Easterbrook, and A. Vyakarnam. 2002. Presence of HIV-1 Gag-specific IFN-gamma+IL-2+ and CD28+IL-2+ CD4 T cell responses is associated with nonprogression in HIV-1 infection. J. Immunol. 169:6376-6385. - PubMed
-
- Brenchley, J. M., B. J. Hill, D. R. Ambrozak, D. A. Price, F. J. Guenaga, J. P. Casazza, J. Kuruppu, J. Yazdani, S. A. Migueles, M. Connors, M. Roederer, D. C. Douek, and R. A. Koup. 2004. T-cell subsets that harbor human immunodeficiency virus (HIV) in vivo: implications for HIV pathogenesis. J. Virol. 78:1160-1168. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
