

Tissue- and tumor-specific targeting of murine leukemia virus-based replication-competent retroviral vectors
- PMID: 16809312
- PMCID: PMC1489065
- DOI: 10.1128/JVI.00020-06
Tissue- and tumor-specific targeting of murine leukemia virus-based replication-competent retroviral vectors
Abstract
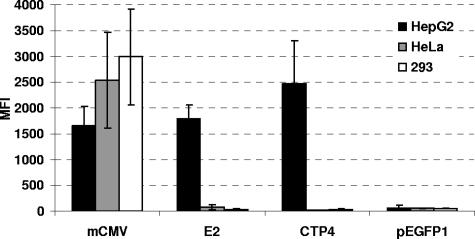
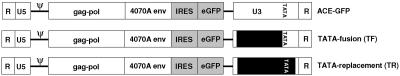
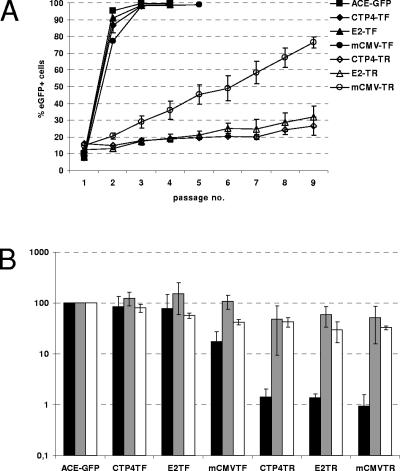
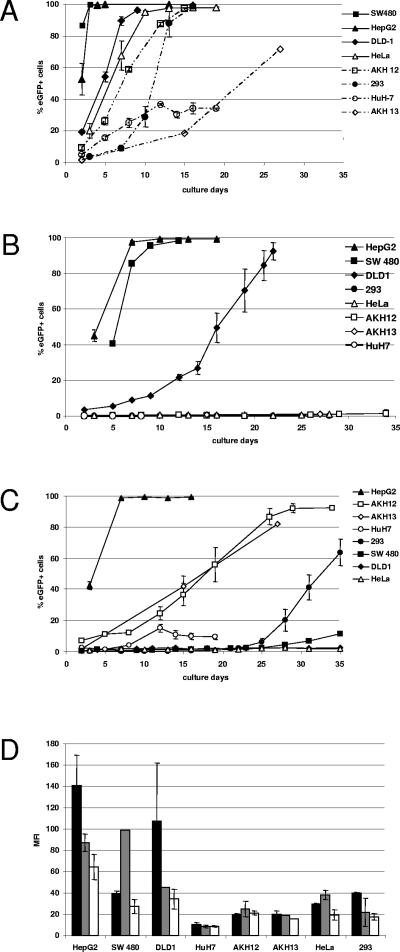
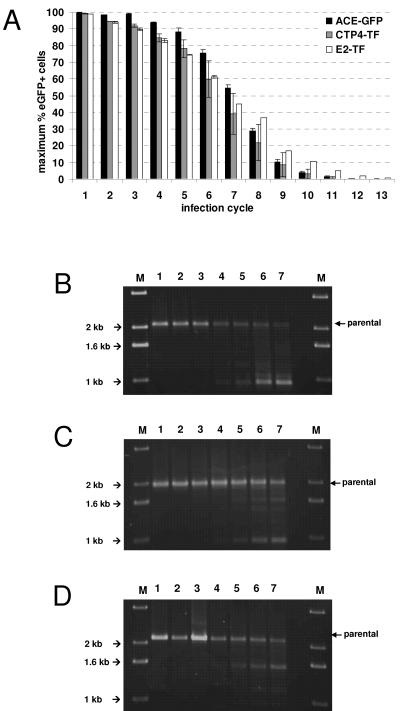
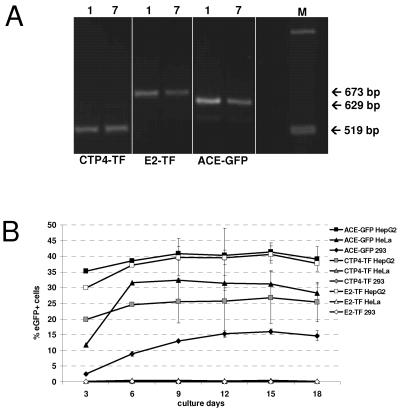
Replication-competent retrovirus vectors based on murine leukemia virus (MLV) have been shown to effectively transfer therapeutic genes over multiple serial infections in cell culture and through solid tumors in vivo with a high degree of genomic stability. While simple retroviruses possess a natural tumor selectivity in that they can transduce only actively dividing cells, additional tumor-targeting strategies would nevertheless be advantageous, since tumor cells are not the only actively dividing cells. In this study, we used the promiscuous murine cytomegalovirus promoter, a chimeric regulatory sequence consisting of the hepatitis B virus enhancer II and the human alpha1-antitrypsin (EII-Pa1AT) promoter, and a synthetic regulatory sequence consisting of a series of T-cell factor binding sites named the CTP4 promoter to generate replicating MLV vectors, whereby the last two are transcriptionally restricted to liver- and beta-catenin/T-cell factor-deregulated cells, respectively. When the heterologous promoters were used to replace almost the entire MLV U3 region, including the MLV TATA box, vector replication was inefficient since nascent virus particle production from infected cells was greatly decreased. Fusion of the heterologous promoters lacking the TATA box to the MLV TATA box, however, generated vectors which replicated with almost-wild-type kinetics throughout permissive cells while exhibiting low or negligible spread in nonpermissive cells. The genomic stability of the vectors was shown to be comparable to that of a similar vector containing wild-type MLV long terminal repeats, and tropism analysis over repeated infection cycles showed that the targeted vectors retained their original specificity.
Figures
Similar articles
-
Tissue-specific transcriptional targeting of a replication-competent retroviral vector.J Virol. 2002 Dec;76(24):12783-91. doi: 10.1128/jvi.76.24.12783-12791.2002. J Virol. 2002. PMID: 12438603 Free PMC article.
-
Antibody-mediated targeting of replication-competent retroviral vectors.Hum Gene Ther. 2003 May 20;14(8):789-802. doi: 10.1089/104303403765255174. Hum Gene Ther. 2003. PMID: 12804141
-
Multiple modifications allow high-titer production of retroviral vectors carrying heterologous regulatory elements.J Virol. 2004 Feb;78(3):1384-92. doi: 10.1128/jvi.78.3.1384-1392.2004. J Virol. 2004. PMID: 14722293 Free PMC article.
-
Beyond oncolytic virotherapy: replication-competent retrovirus vectors for selective and stable transduction of tumors.Curr Gene Ther. 2005 Dec;5(6):655-67. doi: 10.2174/156652305774964659. Curr Gene Ther. 2005. PMID: 16457654 Review.
-
Replication-competent retrovirus vectors for cancer gene therapy.Front Biosci. 2008 Jan 1;13:3083-95. doi: 10.2741/2910. Front Biosci. 2008. PMID: 17981778 Review.
Cited by
-
Human endogenous retroviral long terminal repeat sequences as cell type-specific promoters in retroviral vectors.J Virol. 2009 Dec;83(23):12643-50. doi: 10.1128/JVI.00858-09. Epub 2009 Sep 9. J Virol. 2009. PMID: 19741000 Free PMC article.
-
Tumor-specific suicide gene therapy for hepatocellular carcinoma by transcriptionally targeted retroviral replicating vectors.Gene Ther. 2015 Feb;22(2):155-62. doi: 10.1038/gt.2014.98. Epub 2014 Oct 30. Gene Ther. 2015. PMID: 25354682
-
Endogenous HIV-1 Vpr-mediated apoptosis and proteome alteration of human T-cell leukemia virus-1 transformed C8166 cells.Apoptosis. 2009 Oct;14(10):1212-26. doi: 10.1007/s10495-009-0380-4. Apoptosis. 2009. PMID: 19655254 Free PMC article.
-
Adenovirus-retrovirus hybrid vectors achieve highly enhanced tumor transduction and antitumor efficacy in vivo.Mol Ther. 2011 Jan;19(1):76-82. doi: 10.1038/mt.2010.182. Epub 2010 Aug 31. Mol Ther. 2011. PMID: 20808291 Free PMC article.
-
Molecularly characterised xenograft tumour mouse models: valuable tools for evaluation of new therapeutic strategies for secondary liver cancers.J Biomed Biotechnol. 2009;2009:437284. doi: 10.1155/2009/437284. Epub 2009 Mar 15. J Biomed Biotechnol. 2009. PMID: 19300524 Free PMC article.
References
-
- Addison, C. L., M. Hitt, D. Kunsken, and F. L. Graham. 1997. Comparison of the human versus murine cytomegalovirus immediate early gene promoters for transgene expression by adenoviral vectors. J. Gen. Virol. 78:1653-1661. - PubMed
-
- Boettiger, D., and H. M. Temin. 1970. Light inactivation of focus formation by chicken embryo fibroblasts infected with avian sarcoma virus in the presence of 5-bromodeoxyuridine. Nature 228:622-624. - PubMed
-
- Carruba, G., M. Cervello, M. D. Miceli, R. Farruggio, M. Notarbartolo, L. Virruso, L. Giannitrapani, R. Gambino, G. Montalto, and L. Castagnetta. 1999. Truncated form of beta-catenin and reduced expression of wild-type catenins feature HepG2 human liver cancer cells. Ann. N. Y. Acad. Sci. 886:212-216. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
