

Rac1 links leading edge and uropod events through Rho and myosin activation during chemotaxis
- PMID: 16809619
- PMCID: PMC1895589
- DOI: 10.1182/blood-2006-01-010363
Rac1 links leading edge and uropod events through Rho and myosin activation during chemotaxis
Abstract
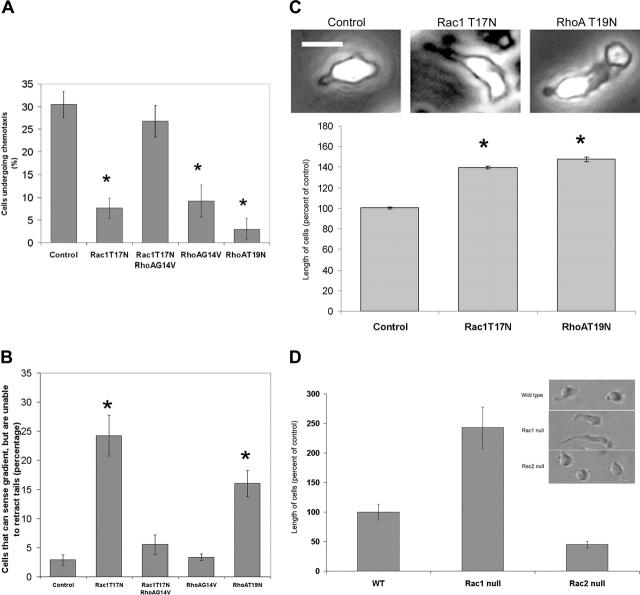
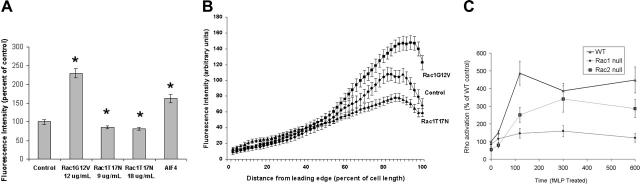
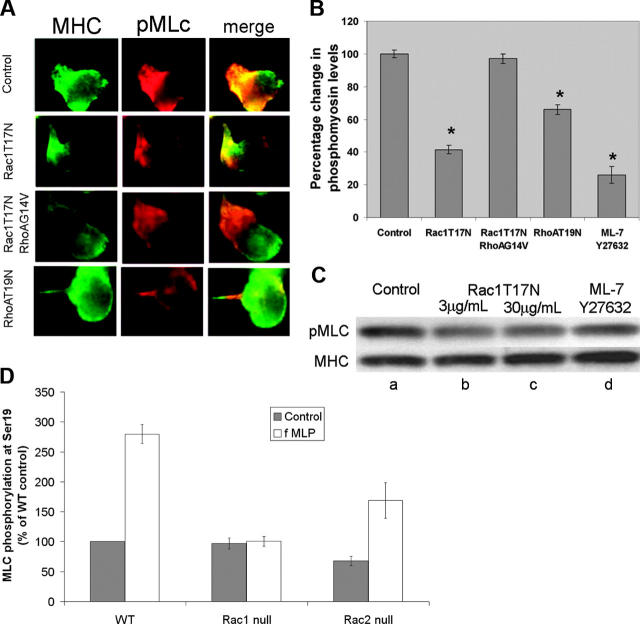
Chemotactic responsiveness is crucial to neutrophil recruitment to sites of infection. During chemotaxis, highly divergent cytoskeletal programs are executed at the leading and trailing edge of motile neutrophils. The Rho family of small GTPases plays a critical role in cell migration, and recent work has focused on elucidating the specific roles played by Rac1, Rac2, Cdc42, and Rho during cellular chemotaxis. Rac GTPases regulate actin polymerization and extension of the leading edge, whereas Rho GTPases control myosin-based contraction of the trailing edge. Rac and Rho signaling are thought to crosstalk with one another, and previous research has focused on mutual inhibition of Rac and Rho signaling during chemotaxis. Indeed, polarization of neutrophils has been proposed to involve the activity of a negative feedback system where Rac activation at the front of the cell inhibits local Rho activation, and vice versa. Using primary human neutrophils and neutrophils derived from a Rac1/Rac2-null transgenic mouse model, we demonstrate here that Rac1 (and not Rac2) is essential for Rho and myosin activation at the trailing edge to regulate uropod function. We conclude that Rac plays both positive and negative roles in the organization of the Rhomyosin "backness" program, thereby promoting stable polarity in chemotaxing neutrophils.
Figures
References
-
- Fenteany G, Glogauer M. Cytoskeletal remodeling in leukocyte function. Curr Opin Hematol. 2004;11: 15-24. - PubMed
-
- Niggli V. Signaling to migration in neutrophils: importance of localized pathways. Int J Biochem Cell Biol. 2003;35: 1619-1638. - PubMed
-
- Niggli V. Rho-kinase in human neutrophils: a role in signalling for myosin light chain phosphorylation and cell migration. FEBS Lett. 1999;445: 69-72. - PubMed
-
- Bokoch GM. Regulation of innate immunity by Rho GTPases. Trends Cell Biol. 2005;15: 163-171. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
