

Oxidation and haem loss kinetics of poly(ethylene glycol)-conjugated haemoglobin (MP4): dissociation between in vitro and in vivo oxidation rates
- PMID: 16813564
- PMCID: PMC1615902
- DOI: 10.1042/BJ20060809
Oxidation and haem loss kinetics of poly(ethylene glycol)-conjugated haemoglobin (MP4): dissociation between in vitro and in vivo oxidation rates
Abstract
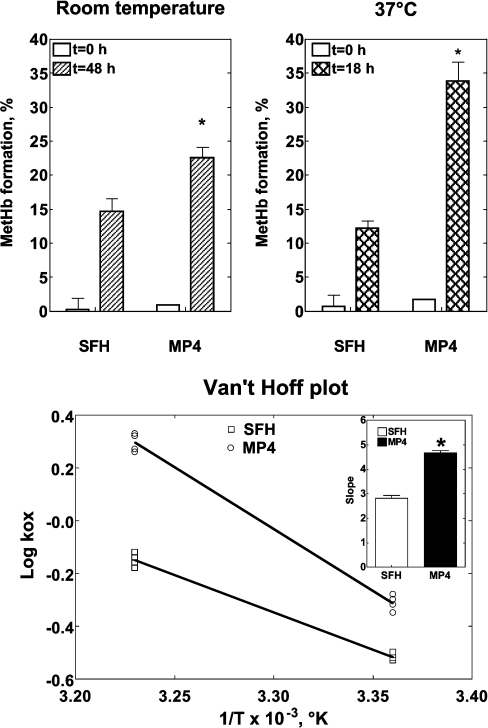
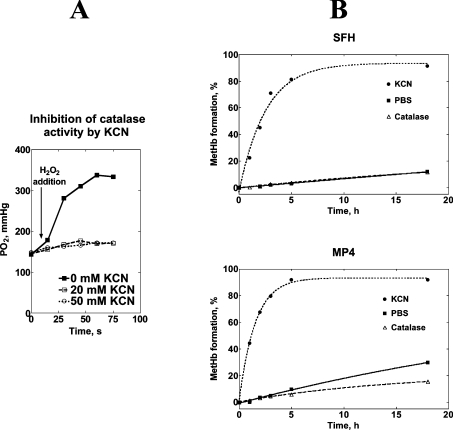
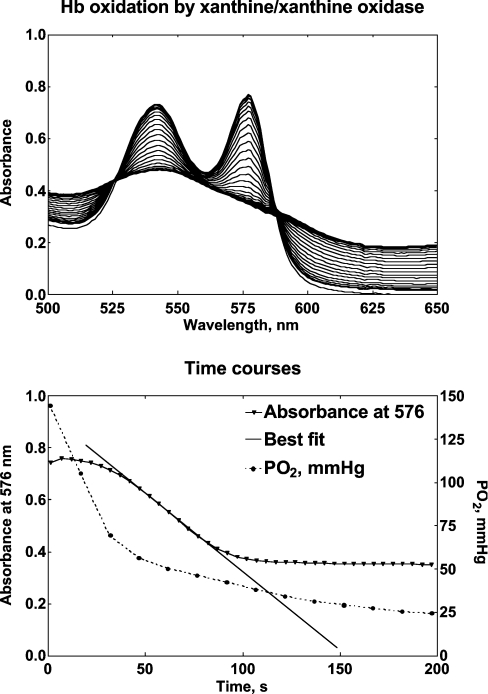
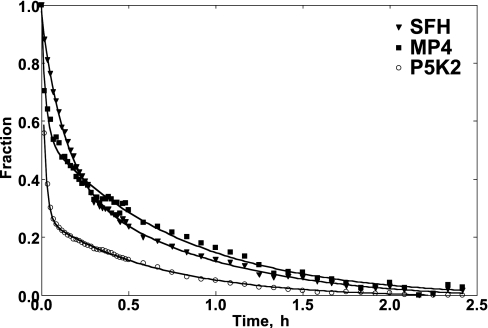
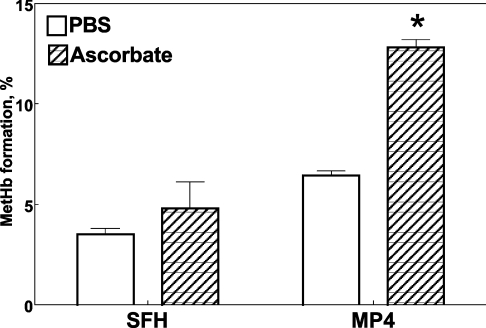
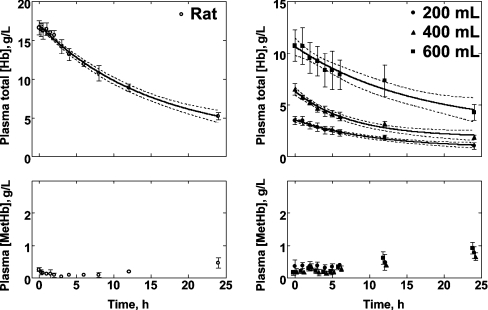
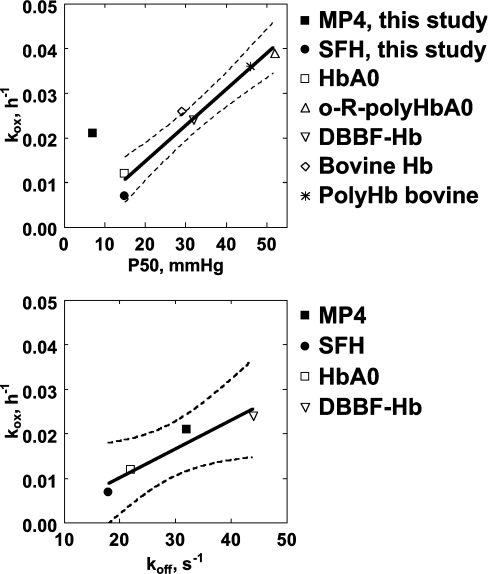
Haemoglobin-based oxygen carriers can undergo oxidation of ferrous haemoglobin into a non-functional ferric form with enhanced rates of haem loss. A recently developed human haemoglobin conjugated to maleimide-activated poly(ethylene glycol), termed MP4, has unique physicochemical properties (increased molecular radius, high oxygen affinity and low cooperativity) and lacks the typical hypertensive response observed with most cell-free haemoglobin solutions. The rate of in vitro MP4 autoxidation is higher compared with the rate for unmodified SFHb (stroma-free haemoglobin), both at room temperature (20-22 degrees C) and at 37 degrees C (P<0.001). This appears to be attributable to residual catalase activity in SFHb but not MP4. In contrast, MP4 and SFHb showed the same susceptibility to oxidation by reactive oxygen species generated by a xanthine-xanthine oxidase system. Once fully oxidized to methaemoglobin, the rate of in vitro haem loss was five times higher in MP4 compared with SFHb in the fast phase, which we assign to the beta subunits, whereas the slow phase (i.e. haem loss from alpha chains) showed similar rates for the two haemoglobins. Formation of MP4 methaemoglobin in vivo following transfusion in rats and humans was slower than predicted by its first-order in vitro autoxidation rate, and there was no appreciable accumulation of MP4 methaemoglobin in plasma before disappearing from the circulation. These results show that MP4 oxidation and haem loss characteristics observed in vitro provide information regarding the effect of poly(ethylene glycol) conjugation on the stability of the haemoglobin molecule, but do not correspond to the oxidation behaviour of MP4 in vivo.
Figures
Similar articles
-
Kinetics of NO and O2 binding to a maleimide poly(ethylene glycol)-conjugated human haemoglobin.Biochem J. 2004 Aug 15;382(Pt 1):183-9. doi: 10.1042/BJ20040156. Biochem J. 2004. PMID: 15175010 Free PMC article.
-
CO-MP4, a polyethylene glycol-conjugated haemoglobin derivative and carbon monoxide carrier that reduces myocardial infarct size in rats.Br J Pharmacol. 2008 Aug;154(8):1649-61. doi: 10.1038/bjp.2008.219. Epub 2008 Jun 9. Br J Pharmacol. 2008. PMID: 18536756 Free PMC article.
-
A phase I single blind clinical trial of a new oxygen transport agent (MP4), human hemoglobin modified with maleimide-activated polyethylene glycol.Haematologica. 2005 Apr;90(4):505-15. Haematologica. 2005. PMID: 15820947 Clinical Trial.
-
Red cell substitutes from hemoglobin--do we start all over again?Curr Opin Chem Biol. 2010 Aug;14(4):538-43. doi: 10.1016/j.cbpa.2010.03.021. Epub 2010 Apr 12. Curr Opin Chem Biol. 2010. PMID: 20392662 Review.
-
Modified haemoglobins as oxygen-transporting blood substitutes.Br J Hosp Med. 1996 Apr 3-16;55(7):443-8. Br J Hosp Med. 1996. PMID: 8730571 Review.
Cited by
-
Biophysical Properties of Lumbricus terrestris Erythrocruorin and Its Potential Use as a Red Blood Cell Substitute.J Funct Biomater. 2012 Jan 6;3(1):49-60. doi: 10.3390/jfb3010049. J Funct Biomater. 2012. PMID: 24956515 Free PMC article.
-
Effects of maleimide-polyethylene glycol-modified human hemoglobin (MP4) on tissue necrosis in SKH1-hr hairless mice.Eur J Med Res. 2009 Mar 17;14(3):123-9. doi: 10.1186/2047-783x-14-3-123. Eur J Med Res. 2009. PMID: 19380283 Free PMC article.
-
PEGylation of αα-Hb using succinimidyl propionic acid PEG 5K: Conjugation chemistry and PEG shell structure dictate respectively the oxygen affinity and resuscitation fluid like properties of PEG αα-Hbs.Artif Cells Nanomed Biotechnol. 2015;43(4):270-81. doi: 10.3109/21691401.2014.885443. Epub 2014 Mar 6. Artif Cells Nanomed Biotechnol. 2015. PMID: 24597567 Free PMC article.
-
Mechanisms of Toxicity and Modulation of Hemoglobin-based Oxygen Carriers.Shock. 2019 Oct;52(1S Suppl 1):41-49. doi: 10.1097/SHK.0000000000001044. Shock. 2019. PMID: 29112106 Free PMC article. Review.
-
Engineering tyrosine residues into hemoglobin enhances heme reduction, decreases oxidative stress and increases vascular retention of a hemoglobin based blood substitute.Free Radic Biol Med. 2019 Apr;134:106-118. doi: 10.1016/j.freeradbiomed.2018.12.030. Epub 2018 Dec 27. Free Radic Biol Med. 2019. PMID: 30594736 Free PMC article.
References
-
- McGown E. L., Lyons M. F., Marini M. A., Zegna A. Reduction of extracellular methemoglobin by erythrocytes. Biochim. Biophys. Acta. 1990;1036:202–206. - PubMed
-
- Reeder B. J., Svistunenko D. A., Cooper C. E., Wilson M. T. The radical and redox chemistry of myoglobin and hemoglobin. Antioxid. Redox Signal. 2004;6:954–966. - PubMed
-
- Motterlini R., Foresti R., Vandegriff K., Intaglietta M., Winslow R. Oxidative-stress response in vascular endothelial cells exposed to acellular hemoglobin solutions. Am. J. Physiol. 1995;269:H648–H655. - PubMed
-
- Brantley R. E., Smerdon S. J., Wilkinson A. J., Singleton E. W., Olson J. S. The mechanism of autooxidation of myoglobin. J. Biol. Chem. 1993;268:6995–7010. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources