

Dopamine presynaptically and heterogeneously modulates nucleus accumbens medium-spiny neuron GABA synapses in vitro
- PMID: 16813648
- PMCID: PMC1538613
- DOI: 10.1186/1471-2202-7-53
Dopamine presynaptically and heterogeneously modulates nucleus accumbens medium-spiny neuron GABA synapses in vitro
Abstract
Background: The striatal complex is the major target of dopamine action in the CNS. There, medium-spiny GABAergic neurons, which constitute about 95% of the neurons in the area, form a mutually inhibitory synaptic network that is modulated by dopamine. When put in culture, the neurons reestablish this network. In particular, they make autaptic connections that provide access to single, identified medium-spiny to medium-spiny neuron synaptic connections.
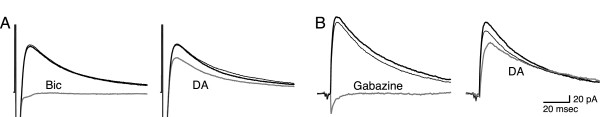
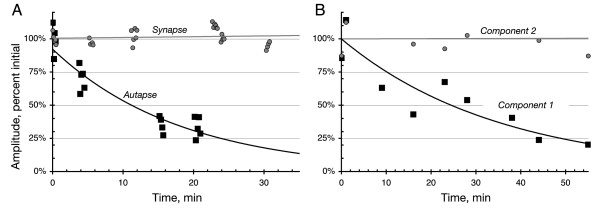
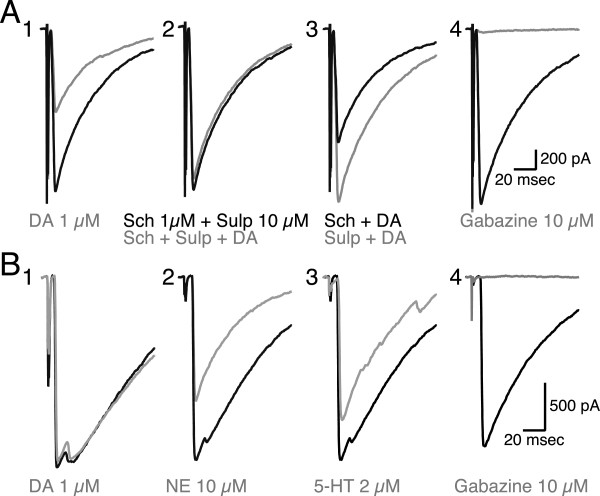
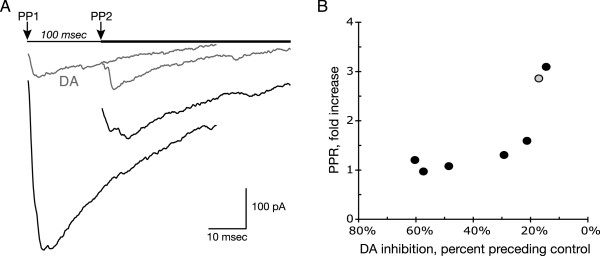
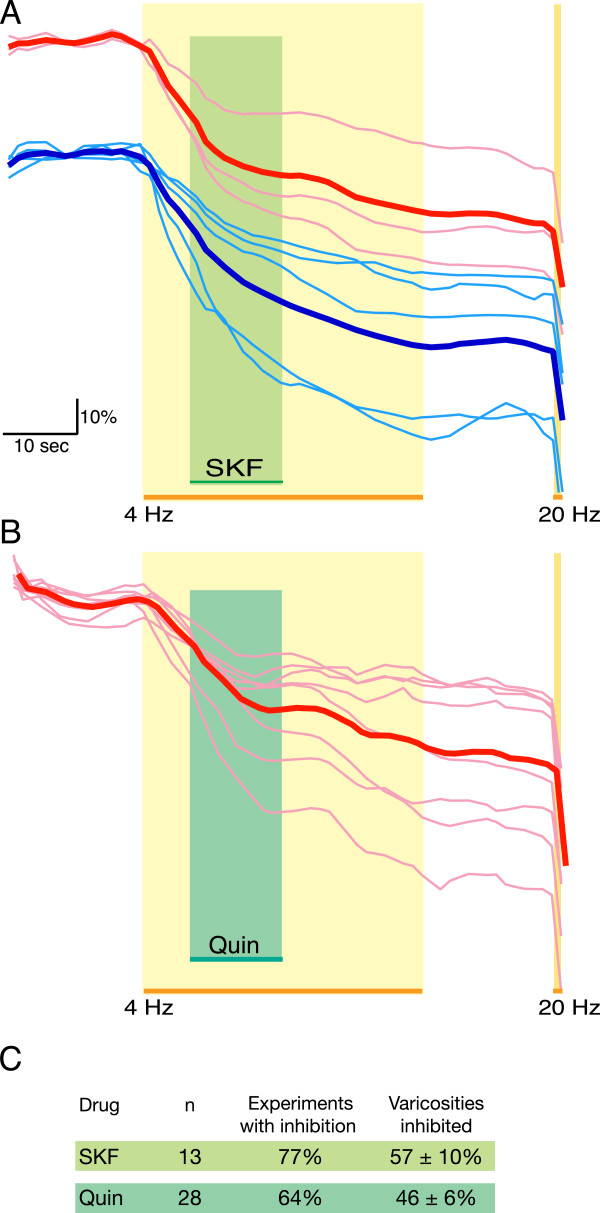
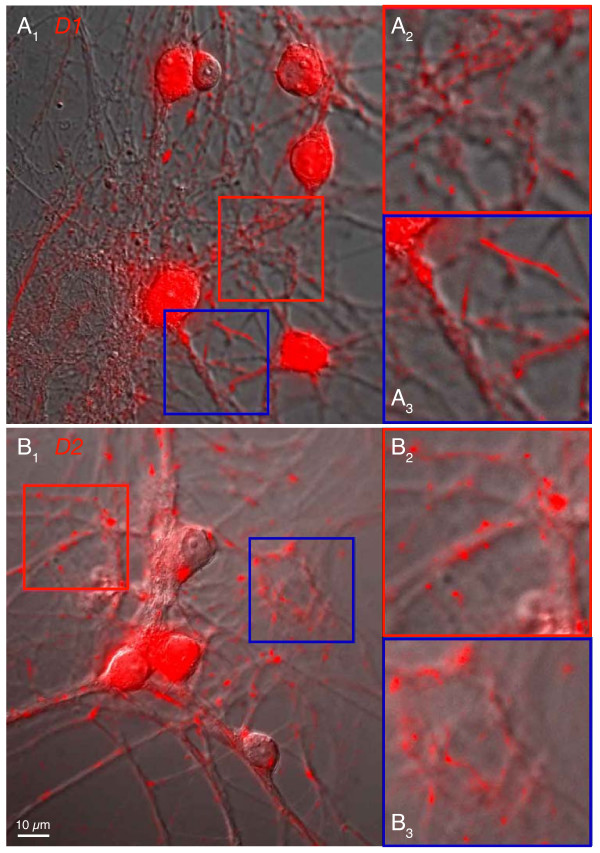
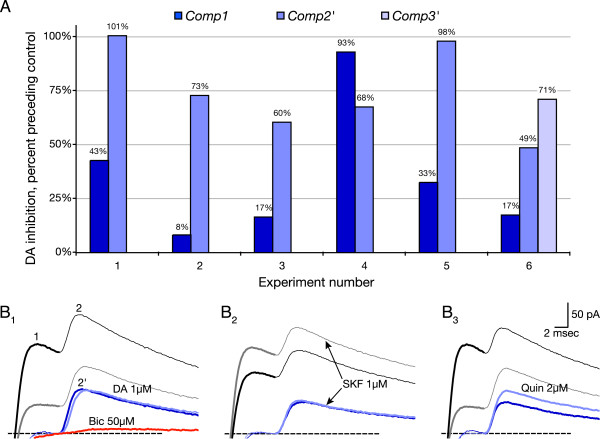
Results: We examined medium-spiny neuron autaptic connections in postnatal cultures from the nucleus accumbens, the ventral part of the striatal complex. These connections were subject to presynaptic dopamine modulation. D1-like receptors mediated either inhibition or facilitation, while D2-like receptors predominantly mediated inhibition. Many connections showed both D1 and D2 modulation, consistent with a significant functional colocalization of D1 and D2-like receptors at presynaptic sites. These same connections were subject to GABAA, GABAB, norepinephrine and serotonin modulation, revealing a multiplicity of modulatory autoreceptors and heteroreceptors on individual varicosities. In some instances, autaptic connections had two components that were differentially modulated by dopamine agonists, suggesting that dopamine receptors could be distributed heterogeneously on the presynaptic varicosities making up a single synaptic (i.e. autaptic) connection.
Conclusion: Differential trafficking of dopamine receptors to different presynaptic varicosities could explain the many controversial studies reporting widely varying degrees of dopamine receptor colocalization in medium-spiny neurons, as well as more generally the diversity of dopamine actions in target areas. Longer-term changes in the modulatory actions of dopamine in the striatal complex could be due to plasticity in the presynaptic distribution of dopamine receptors on medium-spiny neuron varicosities.
Figures
References
-
- Wilson CJ. Basal ganglia. In: Shepherd GM, editor. The Synaptic Organization of the Brain. Fifth. New York , Oxford University Press; 2004. pp. 361–413.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
