

Cooperativity and the origins of rapid, single-exponential kinetics in protein folding
- PMID: 16815915
- PMCID: PMC2242573
- DOI: 10.1110/ps.062180806
Cooperativity and the origins of rapid, single-exponential kinetics in protein folding
Abstract
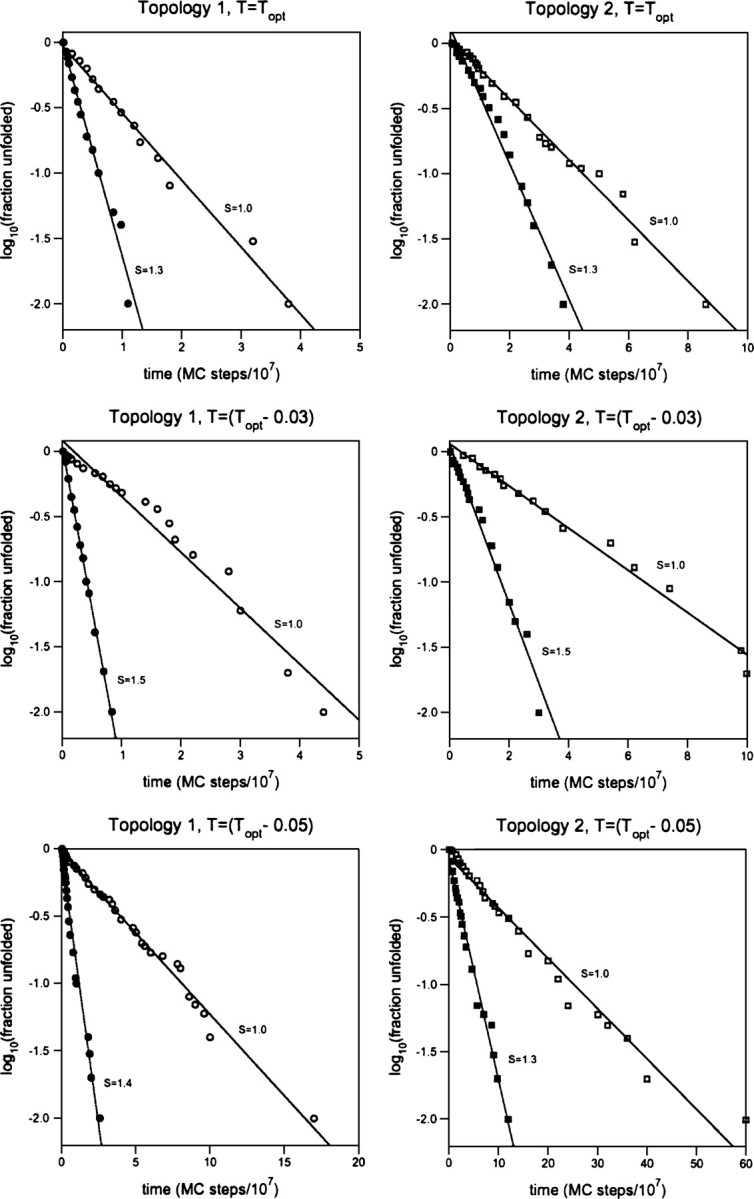
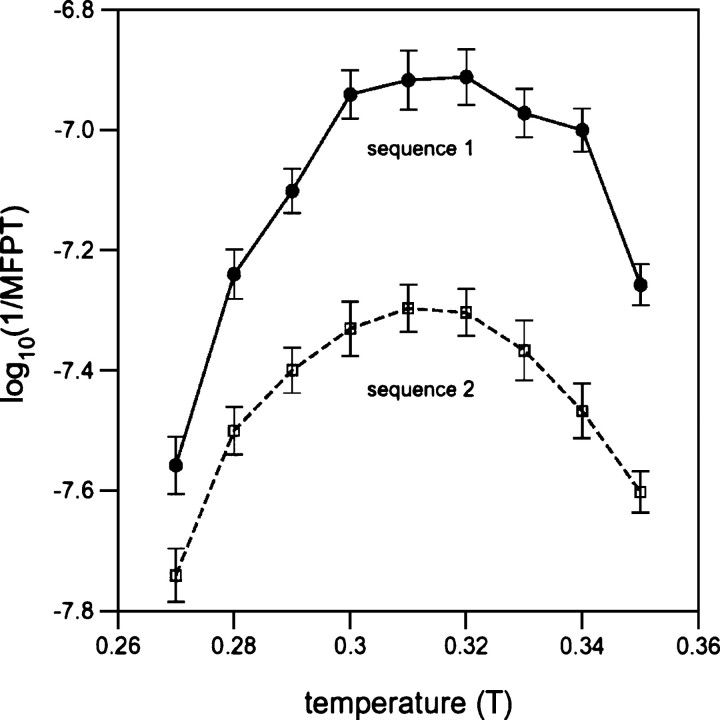
The folding of naturally occurring, single-domain proteins is usually well described as a simple, single-exponential process lacking significant trapped states. Here we further explore the hypothesis that the smooth energy landscape this implies, and the rapid kinetics it engenders, arises due to the extraordinary thermodynamic cooperativity of protein folding. Studying Miyazawa-Jernigan lattice polymers, we find that, even under conditions where the folding energy landscape is relatively optimized (designed sequences folding at their temperature of maximum folding rate), the folding of protein-like heteropolymers is accelerated when their thermodynamic cooperativity is enhanced by enhancing the nonadditivity of their energy potentials. At lower temperatures, where kinetic traps presumably play a more significant role in defining folding rates, we observe still greater cooperativity-induced acceleration. Consistent with these observations, we find that the folding kinetics of our computational models more closely approximates single-exponential behavior as their cooperativity approaches optimal levels. These observations suggest that the rapid folding of naturally occurring proteins is, in part, a consequence of their remarkably cooperative folding.
Figures
Similar articles
-
Cooperativity, smooth energy landscapes and the origins of topology-dependent protein folding rates.J Mol Biol. 2003 Feb 7;326(1):247-53. doi: 10.1016/s0022-2836(02)01356-6. J Mol Biol. 2003. PMID: 12547206
-
The importance of hydration for the kinetics and thermodynamics of protein folding: simplified lattice models.Fold Des. 1998;3(6):523-34. doi: 10.1016/S1359-0278(98)00068-6. Fold Des. 1998. PMID: 9889163
-
Towards a consistent modeling of protein thermodynamic and kinetic cooperativity: how applicable is the transition state picture to folding and unfolding?J Mol Biol. 2002 Jan 25;315(4):899-909. doi: 10.1006/jmbi.2001.5266. J Mol Biol. 2002. PMID: 11812156
-
Cooperativity and protein folding rates.Curr Opin Struct Biol. 2010 Feb;20(1):11-5. doi: 10.1016/j.sbi.2009.12.013. Epub 2010 Jan 21. Curr Opin Struct Biol. 2010. PMID: 20093004 Review.
-
Protein folding in the landscape perspective: chevron plots and non-Arrhenius kinetics.Proteins. 1998 Jan;30(1):2-33. doi: 10.1002/(sici)1097-0134(19980101)30:1<2::aid-prot2>3.0.co;2-r. Proteins. 1998. PMID: 9443337 Review.
Cited by
-
Predicting coupling limits from an experimentally determined energy landscape.Proc Natl Acad Sci U S A. 2007 Mar 20;104(12):4907-12. doi: 10.1073/pnas.0608756104. Epub 2007 Mar 14. Proc Natl Acad Sci U S A. 2007. PMID: 17360387 Free PMC article.
-
Probing the origins of two-state folding.J Chem Phys. 2013 Oct 14;139(14):145104. doi: 10.1063/1.4823502. J Chem Phys. 2013. PMID: 24116650 Free PMC article.
-
Water as a Good Solvent for Unfolded Proteins: Folding and Collapse are Fundamentally Different.J Mol Biol. 2020 Apr 17;432(9):2882-2889. doi: 10.1016/j.jmb.2020.01.031. Epub 2020 Feb 7. J Mol Biol. 2020. PMID: 32044346 Free PMC article. Review.
-
Effective knowledge-based potentials.Protein Sci. 2009 Jul;18(7):1469-85. doi: 10.1002/pro.166. Protein Sci. 2009. PMID: 19530247 Free PMC article.
-
Helix/coil nucleation: a local response to global demands.Biophys J. 2009 Dec 2;97(11):3000-9. doi: 10.1016/j.bpj.2009.09.013. Biophys J. 2009. PMID: 19948130 Free PMC article.
References
-
- Abkevich V.I., Gutin A.M., Shakhnovich E.I. 1995. Impact of local and non-local interactions on thermodynamics and kinetics of protein folding. J. Mol. Biol. 252 460–471. - PubMed
-
- Abkevich V.I., Gutin A.M., Shakhnovich E.I. 1996. Improved design of stable and fast-folding model proteins. Fold. Des. 1 221–230. - PubMed
-
- Bryngelson J.D. and Wolynes P.G. 1989. Intermediates and barrier crossing in a random energy model (with applications to protein folding). J. Phys. Chem. 93 6902–6915.
-
- Bryngelson J.D., Onuchic J.N., Socci N.D., Wolynes P. 1995. Funnels, pathways and the energy landscape of protein folding: A synthesis. Proteins 21 167–195. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
