

Expression and functional analyses of the plastid lipid-associated protein CHRC suggest its role in chromoplastogenesis and stress
- PMID: 16815957
- PMCID: PMC1557619
- DOI: 10.1104/pp.106.082404
Expression and functional analyses of the plastid lipid-associated protein CHRC suggest its role in chromoplastogenesis and stress
Abstract
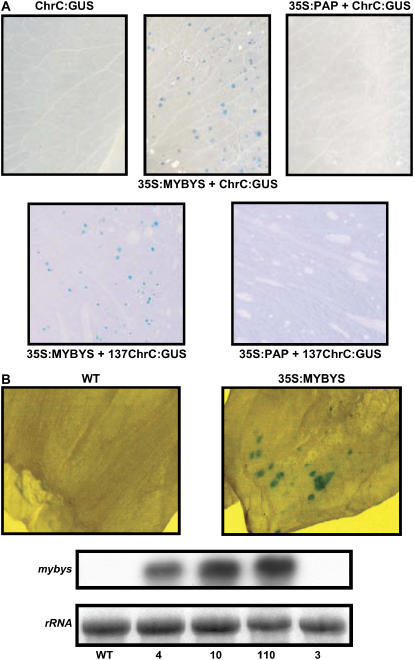
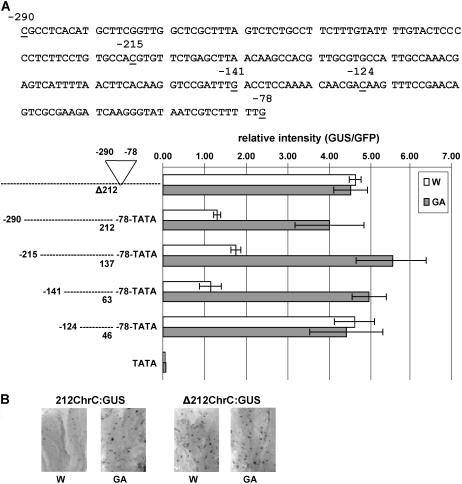
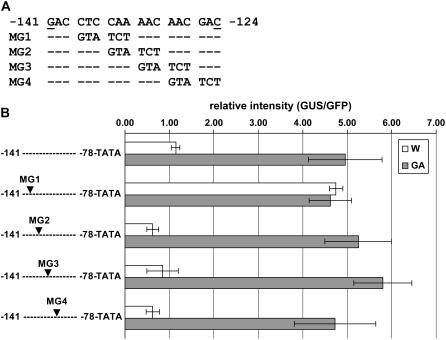
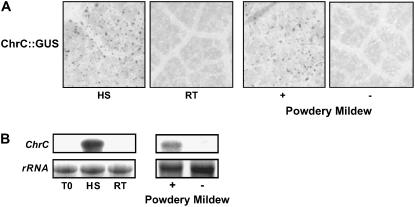
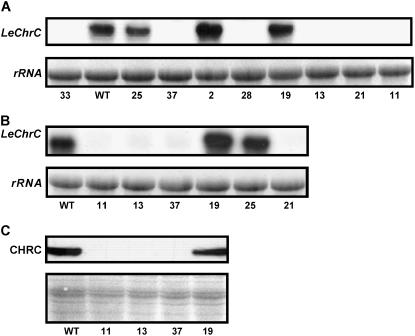
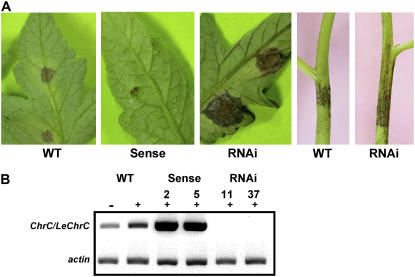
Chromoplastogenesis during flower development and fruit ripening involves the dramatic overaccumulation of carotenoids sequestered into structures containing lipids and proteins called plastid lipid-associated proteins (PAPs). CHRC, a cucumber (Cucumis sativus) PAP, has been suggested to be transcriptionally activated in carotenoid-accumulating flowers by gibberellin (GA). Mybys, a MYB-like trans-activator identified here, may represent a chromoplastogenesis-related factor: Its expression is flower specific and parallels that of ChrC during flower development; moreover, as revealed by stable ectopic and transient-expression assays, it specifically trans-activates ChrC promoter in flowers accumulating carotenoids and flavonoids. A detailed dissection of ChrC promoter revealed a GA-responsive element, gacCTCcaa, the mutation of which abolished ChrC activation by GA. This cis-element is different from the GARE motif and is involved in ChrC activation probably via negative regulation, similar to other GA-responsive systems. The GA responsiveness and MYBYS floral activation of the ChrC promoter do not overlap with respect to cis-elements. To study the functionality of CHRC, which is activated in vegetative tissues similar to other PAPs by various biotic and abiotic stresses, we employed a tomato (Lycopersicon esculentum) plant system and generated RNAi-transgenic lines with suppressed LeCHRC. Transgenic flowers accumulated approximately 30% less carotenoids per unit protein than controls, indicating an interrelationship between PAPs and flower-specific carotenoid accumulation in chromoplasts. Moreover, the transgenic LeCHRC-suppressed plants were significantly more susceptible to Botrytis cinerea infection, suggesting CHRC's involvement in plant protection under stress conditions and supporting the general, evolutionarily preserved role of PAPs.
Figures


Similar articles
-
CHRD, a plant member of the evolutionarily conserved YjgF family, influences photosynthesis and chromoplastogenesis.Planta. 2006 Dec;225(1):89-102. doi: 10.1007/s00425-006-0332-y. Epub 2006 Jul 15. Planta. 2006. PMID: 16845531
-
Molecular cloning of a carotenoid-associated protein from Cucumis sativus corollas: homologous genes involved in carotenoid sequestration in chromoplasts.Plant J. 1996 Dec;10(6):1111-8. doi: 10.1046/j.1365-313x.1996.10061111.x. Plant J. 1996. PMID: 9011091
-
CHRC, encoding a chromoplast-specific carotenoid-associated protein, is an early gibberellic acid-responsive gene.J Biol Chem. 1997 Oct 3;272(40):24747-50. doi: 10.1074/jbc.272.40.24747. J Biol Chem. 1997. PMID: 9312067
-
Chromoplast-specific carotenoid-associated protein appears to be important for enhanced accumulation of carotenoids in hp1 tomato fruits.Plant Physiol. 2013 Apr;161(4):2085-101. doi: 10.1104/pp.112.212191. Epub 2013 Feb 11. Plant Physiol. 2013. PMID: 23400702 Free PMC article.
-
The regulation of carotenoid pigmentation in flowers.Arch Biochem Biophys. 2010 Dec 1;504(1):132-41. doi: 10.1016/j.abb.2010.07.028. Epub 2010 Aug 3. Arch Biochem Biophys. 2010. PMID: 20688043 Review.
Cited by
-
Hydrogen Peroxide Response in Leaves of Poplar (Populus simonii × Populus nigra) Revealed from Physiological and Proteomic Analyses.Int J Mol Sci. 2017 Oct 2;18(10):2085. doi: 10.3390/ijms18102085. Int J Mol Sci. 2017. PMID: 28974034 Free PMC article.
-
Ralstonia solanacearum type III effector RipAF1 mediates plant resistance signaling by ADP-ribosylation of host FBN1.Hortic Res. 2024 Jun 12;11(8):uhae162. doi: 10.1093/hr/uhae162. eCollection 2024 Aug. Hortic Res. 2024. PMID: 39108578 Free PMC article.
-
Transcriptomic changes triggered by carotenoid biosynthesis inhibitors and role of Citrus sinensis phosphate transporter 4;2 (CsPHT4;2) in enhancing carotenoid accumulation.Planta. 2019 Jan;249(1):257-270. doi: 10.1007/s00425-018-2970-2. Epub 2018 Aug 6. Planta. 2019. PMID: 30083809
-
The R2R3-MYB transcription factor EVER controls the emission of petunia floral volatiles by regulating epicuticular wax biosynthesis in the petal epidermis.Plant Cell. 2023 Dec 21;36(1):174-193. doi: 10.1093/plcell/koad251. Plant Cell. 2023. PMID: 37818992 Free PMC article.
-
Carotenoid metabolism and regulation in horticultural crops.Hortic Res. 2015 Aug 26;2:15036. doi: 10.1038/hortres.2015.36. eCollection 2015. Hortic Res. 2015. PMID: 26504578 Free PMC article. Review.
References
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K (2001) Current Protocols in Molecular Biology. John Wiley and Sons, New York
-
- Ben-Meir H (2003) Characterization of the flavonoid biosynthetic pathway in Gypsophila flowers. PhD thesis. Hebrew University of Jerusalem, Jerusalem
-
- Ben-Nissan G, Lee J-Y, Borohov A, Weiss D (2004) GIP, a Petunia hybrida GA-induced cysteine-rich protein: a possible role in shoot elongation and transition to flowering. Plant J 37: 229–238 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
