

A variable cluster of ethylene response factor-like genes regulates metabolic and developmental acclimation responses to submergence in rice
- PMID: 16816135
- PMCID: PMC1533987
- DOI: 10.1105/tpc.106.043000
A variable cluster of ethylene response factor-like genes regulates metabolic and developmental acclimation responses to submergence in rice
Abstract
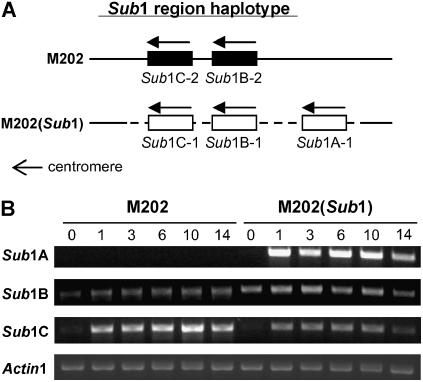
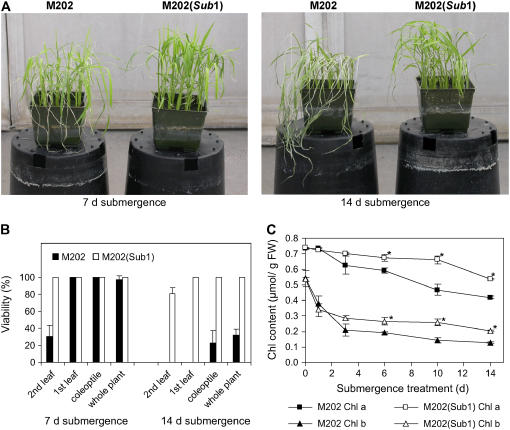
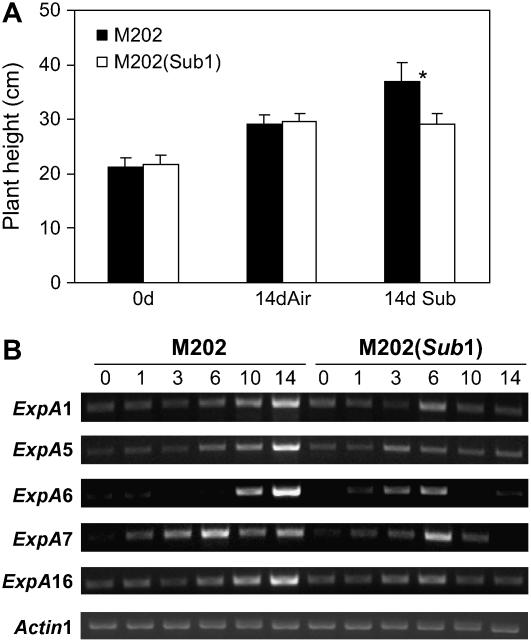
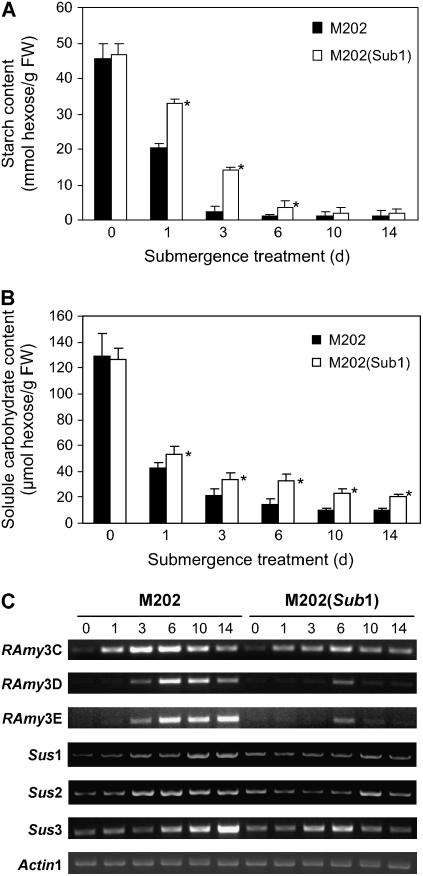
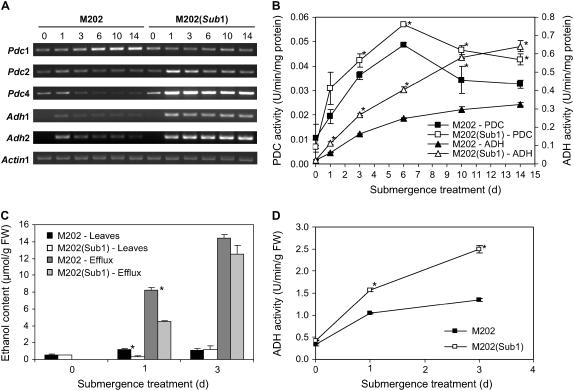
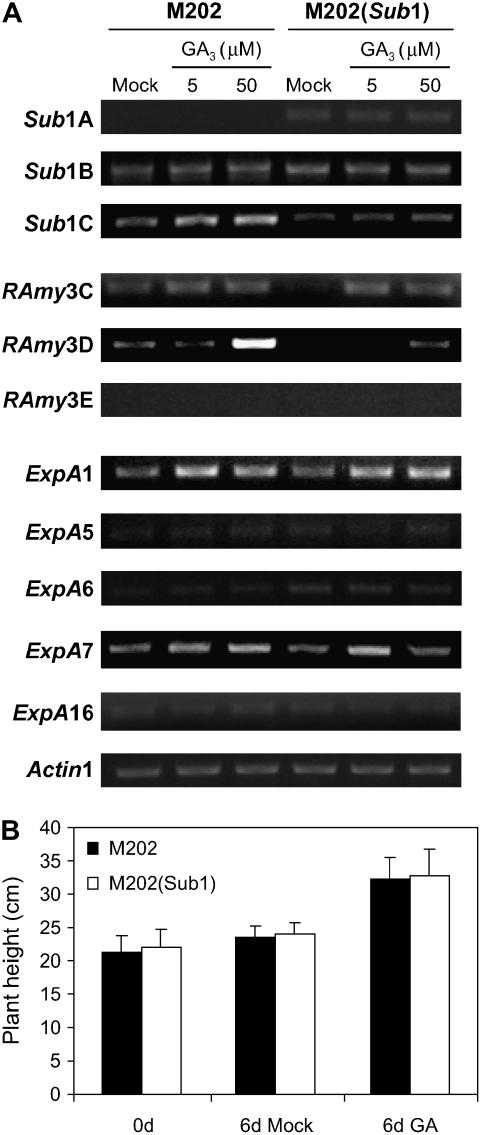
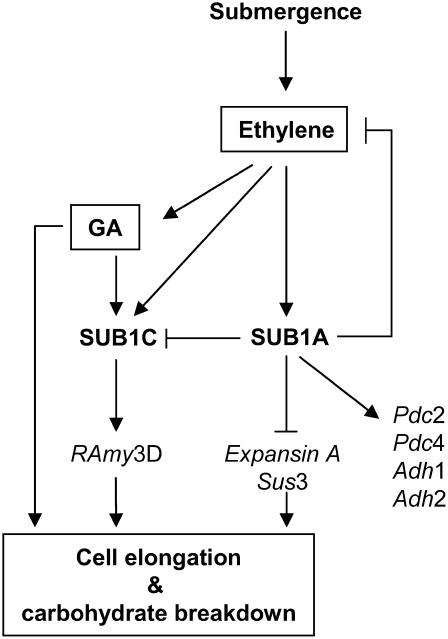
Submergence-1 (Sub1), a major quantitative trait locus affecting tolerance to complete submergence in lowland rice (Oryza sativa), contains two or three ethylene response factor (ERF)-like genes whose transcripts are regulated by submergence. In the submergence-intolerant japonica cultivar M202, this locus encodes two ERF genes, Sub1B and Sub1C. In the tolerant near-isogenic line containing the Sub1 locus from the indica FR13A, M202(Sub1), the locus additionally encodes the ERF gene Sub1A. During submergence, the tolerant M202(Sub1) displayed restrained leaf and internode elongation, chlorophyll degradation, and carbohydrate consumption, whereas the enzymatic activities of pyruvate decarboxylase and alcohol dehydrogenase were increased significantly compared with the intolerant M202. Transcript levels of genes associated with carbohydrate consumption, ethanolic fermentation, and cell expansion were distinctly regulated in the two lines. Sub1A and Sub1C transcript levels were shown to be upregulated by submergence and ethylene, with the Sub1C allele in M202 also upregulated by treatment with gibberellic acid (GA). These findings demonstrate that the Sub1 region haplotype determines ethylene- and GA-mediated metabolic and developmental responses to submergence through differential expression of Sub1A and Sub1C. Submergence tolerance in lowland rice is conferred by a specific allele variant of Sub1A that dampens ethylene production and GA responsiveness, causing quiescence in growth that correlates with the capacity for regrowth upon desubmergence.
Figures

Similar articles
-
Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice.Nature. 2006 Aug 10;442(7103):705-8. doi: 10.1038/nature04920. Nature. 2006. PMID: 16900200
-
Expression of rice SUB1A and SUB1C transcription factors in Arabidopsis uncovers flowering inhibition as a submergence tolerance mechanism.Plant J. 2011 Aug;67(3):434-46. doi: 10.1111/j.1365-313X.2011.04605.x. Epub 2011 Jun 14. Plant J. 2011. PMID: 21481028
-
The submergence tolerance regulator Sub1A mediates stress-responsive expression of AP2/ERF transcription factors.Plant Physiol. 2010 Mar;152(3):1674-92. doi: 10.1104/pp.109.152157. Epub 2010 Jan 27. Plant Physiol. 2010. PMID: 20107022 Free PMC article.
-
[Submergence tolerance and Sub1 locus in rice].Yi Chuan. 2010 Sep;32(9):886-92. Yi Chuan. 2010. PMID: 20870609 Review. Chinese.
-
Unraveling the genetic enigma of rice submergence tolerance: Shedding light on the role of ethylene response factor-encoding gene SUB1A-1.Plant Physiol Biochem. 2024 Jan;206:108224. doi: 10.1016/j.plaphy.2023.108224. Epub 2023 Nov 25. Plant Physiol Biochem. 2024. PMID: 38091930 Review.
Cited by
-
The submergence tolerance gene SUB1A delays leaf senescence under prolonged darkness through hormonal regulation in rice.Plant Physiol. 2012 Dec;160(4):1795-807. doi: 10.1104/pp.112.207738. Epub 2012 Oct 16. Plant Physiol. 2012. PMID: 23073696 Free PMC article.
-
Ethylene-Mediated Acclimations to Flooding Stress.Plant Physiol. 2015 Sep;169(1):3-12. doi: 10.1104/pp.15.00387. Epub 2015 Apr 20. Plant Physiol. 2015. PMID: 25897003 Free PMC article. Review.
-
Endogenous abscisic acid as a key switch for natural variation in flooding-induced shoot elongation.Plant Physiol. 2010 Oct;154(2):969-77. doi: 10.1104/pp.110.162792. Epub 2010 Aug 10. Plant Physiol. 2010. PMID: 20699400 Free PMC article.
-
ERF Transcription Factor OsBIERF3 Positively Contributes to Immunity against Fungal and Bacterial Diseases but Negatively Regulates Cold Tolerance in Rice.Int J Mol Sci. 2022 Jan 6;23(2):606. doi: 10.3390/ijms23020606. Int J Mol Sci. 2022. PMID: 35054806 Free PMC article.
-
Differential molecular responses of rice and wheat coleoptiles to anoxia reveal novel metabolic adaptations in amino acid metabolism for tissue tolerance.Plant Physiol. 2011 Aug;156(4):1706-24. doi: 10.1104/pp.111.175570. Epub 2011 May 27. Plant Physiol. 2011. PMID: 21622811 Free PMC article.
References
-
- Armstrong, W., and Drew, M.C. (2002). Root growth and metabolism under oxygen deficiency. In Plant Roots: The Hidden Half, 3rd ed. Y. Waisel, A. Eshel, and U. Kafkafi, eds (New York: Marcel Dekker), pp. 729–761.
-
- Benschop, J.J., Jackson, M.B., Gühl, K., Vreeburg, R.A.M., Croker, S.J., Peeters, A.J.M., and Voesenek, A.C.J. (2005). Contrasting interactions between ethylene and abscisic acid in Rumex species differing in submergence tolerance. Plant J. 44 756–768. - PubMed
-
- Bethke, P.C., Schuurink, R., and Jones, R.L. (1997). Hormonal signaling in cereal aleurone. J. Exp. Bot. 48 1337–1356.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
