

Limited role for the DsrA and RprA regulatory RNAs in rpoS regulation in Salmonella enterica
- PMID: 16816180
- PMCID: PMC1539969
- DOI: 10.1128/JB.00206-06
Limited role for the DsrA and RprA regulatory RNAs in rpoS regulation in Salmonella enterica
Abstract
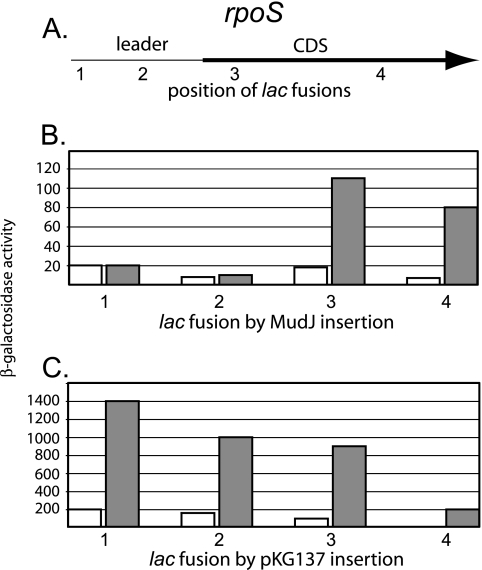
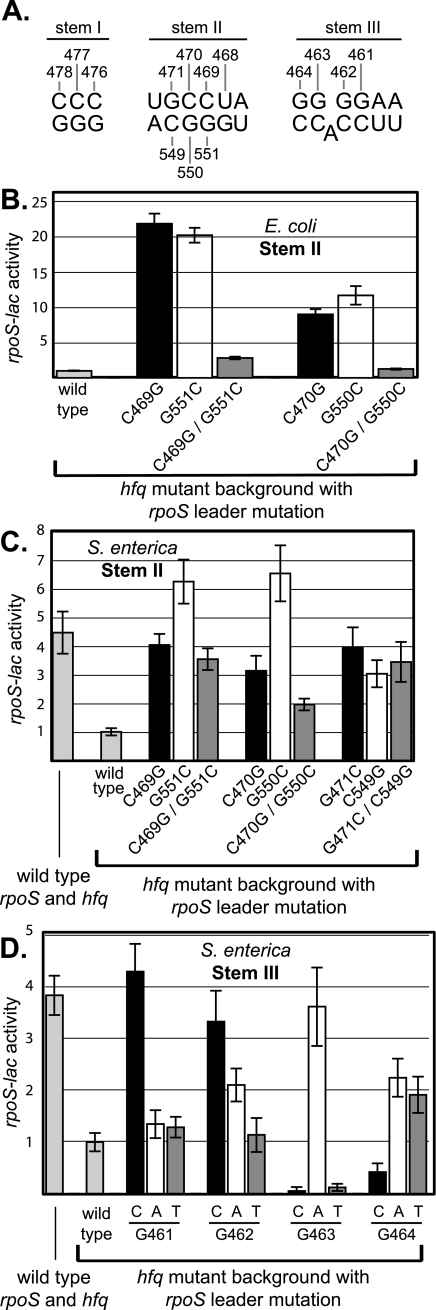
RpoS, the sigma factor of enteric bacteria that responds to stress and stationary phase, is subject to complex regulation acting at multiple levels, including transcription, translation, and proteolysis. Increased translation of rpoS mRNA during growth at low temperature, after osmotic challenge, or with a constitutively activated Rcs phosphorelay depends on two trans-acting small regulatory RNAs (sRNAs) in Escherichia coli. The DsrA and RprA sRNAs are both highly conserved in Salmonella enterica, as is their target, an inhibitory antisense element within the rpoS untranslated leader. Analysis of dsrA and rprA deletion mutants indicates that while the increased translation of RpoS in response to osmotic challenge is conserved in S. enterica, dependence on these two sRNA regulators is much reduced. Furthermore, low-temperature growth or constitutive RcsC activation had only modest effects on RpoS expression, and these increases were, respectively, independent of dsrA or rprA function. This lack of conservation of sRNA function suggests surprising flexibility in RpoS regulation.
Figures





Similar articles
-
Regulation of RpoS by a novel small RNA: the characterization of RprA.Mol Microbiol. 2001 Mar;39(5):1382-94. doi: 10.1111/j.1365-2958.2001.02329.x. Mol Microbiol. 2001. PMID: 11251852
-
Mechanism of positive regulation by DsrA and RprA small noncoding RNAs: pairing increases translation and protects rpoS mRNA from degradation.J Bacteriol. 2010 Nov;192(21):5559-71. doi: 10.1128/JB.00464-10. Epub 2010 Aug 27. J Bacteriol. 2010. PMID: 20802038 Free PMC article.
-
Effect of Hfq on RprA-rpoS mRNA pairing: Hfq-RNA binding and the influence of the 5' rpoS mRNA leader region.Biochemistry. 2008 Oct 28;47(43):11184-95. doi: 10.1021/bi800479p. Epub 2008 Oct 1. Biochemistry. 2008. PMID: 18826256
-
Small non-coding RNAs, co-ordinators of adaptation processes in Escherichia coli: the RpoS paradigm.Mol Microbiol. 2003 May;48(4):855-61. doi: 10.1046/j.1365-2958.2003.03454.x. Mol Microbiol. 2003. PMID: 12753181 Review.
-
Riboregulation by DsrA RNA: trans-actions for global economy.Mol Microbiol. 2000 Nov;38(4):667-72. doi: 10.1046/j.1365-2958.2000.02162.x. Mol Microbiol. 2000. PMID: 11115103 Review.
Cited by
-
Effect of salt and RNA structure on annealing and strand displacement by Hfq.Nucleic Acids Res. 2009 Oct;37(18):6205-13. doi: 10.1093/nar/gkp646. Epub 2009 Aug 11. Nucleic Acids Res. 2009. PMID: 19671524 Free PMC article.
-
A novel insight on signal transduction mechanism of RcsCDB system in Salmonella enterica serovar typhimurium.PLoS One. 2013 Sep 4;8(9):e72527. doi: 10.1371/journal.pone.0072527. eCollection 2013. PLoS One. 2013. PMID: 24023746 Free PMC article.
-
Deep sequencing analysis of small noncoding RNA and mRNA targets of the global post-transcriptional regulator, Hfq.PLoS Genet. 2008 Aug 22;4(8):e1000163. doi: 10.1371/journal.pgen.1000163. PLoS Genet. 2008. PMID: 18725932 Free PMC article.
-
The RNA chaperone Hfq is essential for the virulence of Salmonella typhimurium.Mol Microbiol. 2007 Jan;63(1):193-217. doi: 10.1111/j.1365-2958.2006.05489.x. Epub 2006 Dec 5. Mol Microbiol. 2007. PMID: 17163975 Free PMC article.
-
Small RNA-based feedforward loop with AND-gate logic regulates extrachromosomal DNA transfer in Salmonella.Proc Natl Acad Sci U S A. 2015 Aug 25;112(34):E4772-81. doi: 10.1073/pnas.1507825112. Epub 2015 Aug 11. Proc Natl Acad Sci U S A. 2015. PMID: 26307765 Free PMC article.
References
-
- Altman, E., J. R. Roth, A. Hessel, and K. E. Sanderson. 1996. Transposons currently in use in genetic analysis of Salmonella species, p. 2613-2626. In F. C. Neidhardt, R. Curtiss III, J. L. Ingraham, E. C. C. Lin, K. B. Low, B. Magasanik, W. S. Reznikoff, M. Riley, M. Schaechter, and H. E. Umbarger (ed.), Escherichia coli and Salmonella: cellular and molecular biology, 2nd ed., vol. 2. American Society for Microbiology, Washington, D.C.
-
- Bochner, B. R., and B. N. Ames. 1982. Complete analysis of cellular nucleotides by two-dimensional thin layer chromatography. J. Biol. Chem. 257:9759-9769. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
