

Genetic exchange between homeologous sequences in mammalian chromosomes is averted by local homology requirements for initiation and resolution of recombination
- PMID: 16816418
- PMCID: PMC1569803
- DOI: 10.1534/genetics.106.060590
Genetic exchange between homeologous sequences in mammalian chromosomes is averted by local homology requirements for initiation and resolution of recombination
Abstract
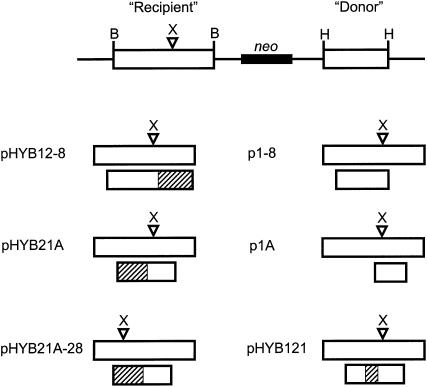
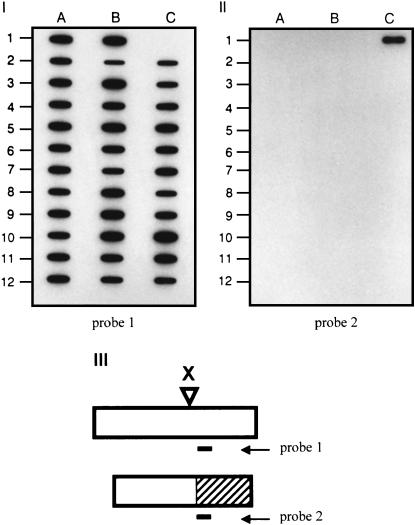
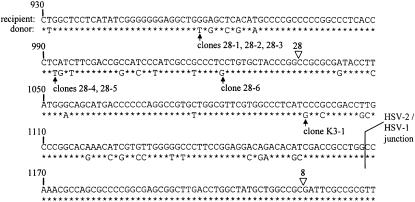
We examined the mechanism by which recombination between imperfectly matched sequences (homeologous recombination) is suppressed in mammalian chromosomes. DNA substrates were constructed, each containing a thymidine kinase (tk) gene disrupted by insertion of an XhoI linker and referred to as a "recipient" gene. Each substrate also contained one of several "donor" tk sequences that could potentially correct the recipient gene via recombination. Each donor sequence either was perfectly homologous to the recipient gene or contained homeologous sequence sharing only 80% identity with the recipient gene. Mouse Ltk(-) fibroblasts were stably transfected with the various substrates and tk(+) segregants produced via intrachromosomal recombination were recovered. We observed exclusion of homeologous sequence from gene conversion tracts when homeologous sequence was positioned adjacent to homologous sequence in the donor but not when homeologous sequence was surrounded by homology in the donor. Our results support a model in which homeologous recombination in mammalian chromosomes is suppressed by a nondestructive dismantling of mismatched heteroduplex DNA (hDNA) intermediates. We suggest that mammalian cells do not dismantle mismatched hDNA by responding to mismatches in hDNA per se but rather rejection of mismatched hDNA appears to be driven by a requirement for localized homology for resolution of recombination.
Figures
Similar articles
-
Fine-resolution analysis of products of intrachromosomal homeologous recombination in mammalian cells.Mol Cell Biol. 1997 Jul;17(7):3614-28. doi: 10.1128/MCB.17.7.3614. Mol Cell Biol. 1997. PMID: 9199296 Free PMC article.
-
High homology is not required at the site of strand invasion during recombinational double-strand break repair in mammalian chromosomes.DNA Repair (Amst). 2017 Dec;60:1-8. doi: 10.1016/j.dnarep.2017.10.006. Epub 2017 Oct 16. DNA Repair (Amst). 2017. PMID: 29055804
-
Mismatch tolerance during homologous recombination in mammalian cells.DNA Repair (Amst). 2018 Oct;70:25-36. doi: 10.1016/j.dnarep.2018.07.011. Epub 2018 Aug 1. DNA Repair (Amst). 2018. PMID: 30103093
-
Ensuring the fidelity of recombination in mammalian chromosomes.Bioessays. 2008 Nov;30(11-12):1163-71. doi: 10.1002/bies.20845. Bioessays. 2008. PMID: 18937366 Review.
-
Mismatch repair and homeologous recombination.DNA Repair (Amst). 2016 Feb;38:75-83. doi: 10.1016/j.dnarep.2015.11.010. Epub 2015 Dec 7. DNA Repair (Amst). 2016. PMID: 26739221 Review.
Cited by
-
Mechanisms of ectopic gene conversion.Genes (Basel). 2010 Nov 29;1(3):427-39. doi: 10.3390/genes1030427. Genes (Basel). 2010. PMID: 24000309 Free PMC article.
-
Accurate homologous recombination is a prominent double-strand break repair pathway in mammalian chromosomes and is modulated by mismatch repair protein Msh2.Mol Cell Biol. 2007 Nov;27(22):7816-27. doi: 10.1128/MCB.00455-07. Epub 2007 Sep 10. Mol Cell Biol. 2007. PMID: 17846123 Free PMC article.
-
The Rate and Tract Length of Gene Conversion between Duplicated Genes.Genes (Basel). 2011 Mar 25;2(2):313-31. doi: 10.3390/genes2020313. Genes (Basel). 2011. PMID: 24710193 Free PMC article.
-
Induction of recombination between diverged sequences in a mammalian genome by a double-strand break.Cell Mol Life Sci. 2014 Jun;71(12):2359-71. doi: 10.1007/s00018-013-1520-0. Epub 2013 Nov 21. Cell Mol Life Sci. 2014. PMID: 24257896 Free PMC article.
-
Plant mitochondrial recombination surveillance requires unusual RecA and MutS homologs.Plant Cell. 2007 Apr;19(4):1251-64. doi: 10.1105/tpc.106.048355. Epub 2007 Apr 27. Plant Cell. 2007. PMID: 17468263 Free PMC article.
References
-
- Bacolla, A., and R. D. Wells, 2004. Non-B DNA conformations, genomic rearrangements, and human disease. J. Biol. Chem. 279: 47411–47414. - PubMed
-
- Bazemore, L. R., M. Takahasgi and C. M. Radding, 1997. Kinetic analysis of pairing and strand exchange catalyzed by RecA. J. Biol. Chem. 272: 14672–14682. - PubMed
-
- Braybrooke, J. P., J. L. Li, L. Wu, F. Caple, F. E. Benson et al., 2003. Functional interaction between the Bloom's syndrome helicase and the RAD51 paralog, RAD51L3 (RAD51D). J. Biol. Chem. 278: 48357–48366. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
