

A genomewide screen for suppressors of par-2 uncovers potential regulators of PAR protein-dependent cell polarity in Caenorhabditis elegans
- PMID: 16816419
- PMCID: PMC1569778
- DOI: 10.1534/genetics.106.060517
A genomewide screen for suppressors of par-2 uncovers potential regulators of PAR protein-dependent cell polarity in Caenorhabditis elegans
Abstract
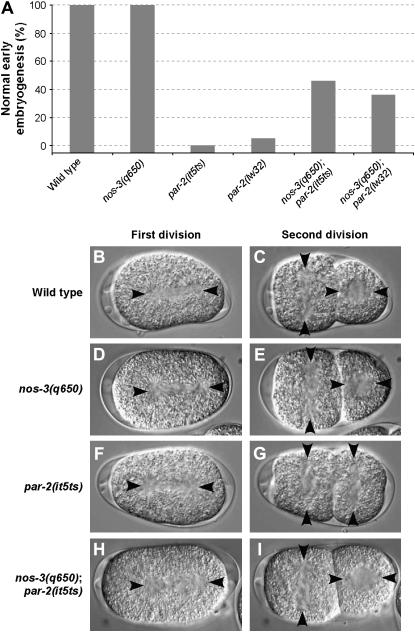
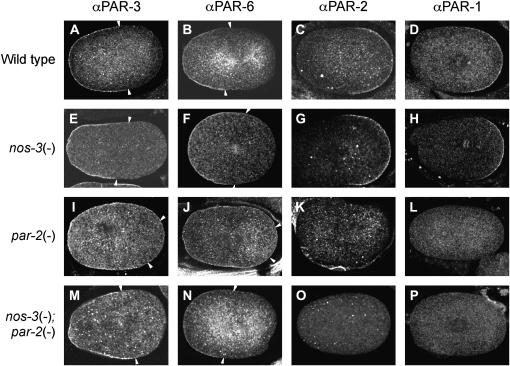
The PAR proteins play an essential role in establishing and maintaining cell polarity. While their function is conserved across species, little is known about their regulators and effectors. Here we report the identification of 13 potential components of the C. elegans PAR polarity pathway, identified in an RNAi-based, systematic screen to find suppressors of par-2(it5ts) lethality. Most of these genes are conserved in other species. Phenotypic analysis of double-mutant animals revealed that some of the suppressors can suppress lethality associated with the strong loss-of-function allele par-2(lw32), indicating that they might impinge on the PAR pathway independently of the PAR-2 protein. One of these is the gene nos-3, which encodes a homolog of Drosophila Nanos. We find that nos-3 suppresses most of the phenotypes associated with loss of par-2 function, including early cell division defects and maternal-effect sterility. Strikingly, while PAR-1 activity was essential in nos-3; par-2 double mutants, its asymmetric localization at the posterior cortex was not restored, suggesting that the function of PAR-1 is independent of its cortical localization. Taken together, our results identify conserved components that regulate PAR protein function and also suggest a role for NOS-3 in PAR protein-dependent cell polarity.
Figures

Similar articles
-
PAR-6 levels are regulated by NOS-3 in a CUL-2 dependent manner in Caenorhabditiselegans.Dev Biol. 2008 Jul 15;319(2):267-72. doi: 10.1016/j.ydbio.2008.04.016. Epub 2008 Apr 23. Dev Biol. 2008. PMID: 18502413
-
CDC-42 and RHO-1 coordinate acto-myosin contractility and PAR protein localization during polarity establishment in C. elegans embryos.Development. 2006 Sep;133(18):3507-16. doi: 10.1242/dev.02527. Epub 2006 Aug 9. Development. 2006. PMID: 16899536
-
Pod-2, along with pod-1, defines a new class of genes required for polarity in the early Caenorhabditis elegans embryo.Dev Biol. 2001 May 15;233(2):412-24. doi: 10.1006/dbio.2001.0234. Dev Biol. 2001. PMID: 11336504
-
Cell polarity and asymmetric cell division: the C. elegans early embryo.Essays Biochem. 2012;53:1-14. doi: 10.1042/bse0530001. Essays Biochem. 2012. PMID: 22928504 Review.
-
Anterior-posterior polarity in C. elegans and Drosophila--PARallels and differences.Science. 2002 Dec 6;298(5600):1946-50. doi: 10.1126/science.1072162. Science. 2002. PMID: 12471246 Review.
Cited by
-
The PAR network: redundancy and robustness in a symmetry-breaking system.Philos Trans R Soc Lond B Biol Sci. 2013 Sep 23;368(1629):20130010. doi: 10.1098/rstb.2013.0010. Print 2013. Philos Trans R Soc Lond B Biol Sci. 2013. PMID: 24062581 Free PMC article. Review.
-
The identification of inhibitors of Schistosoma mansoni miracidial transformation by incorporating a medium-throughput small-molecule screen.Exp Parasitol. 2010 Jun;125(2):84-94. doi: 10.1016/j.exppara.2009.12.021. Epub 2010 Jan 11. Exp Parasitol. 2010. PMID: 20060828 Free PMC article.
-
Systematic genetic interaction screens uncover cell polarity regulators and functional redundancy.Nat Cell Biol. 2013 Jan;15(1):103-12. doi: 10.1038/ncb2639. Epub 2012 Dec 16. Nat Cell Biol. 2013. PMID: 23242217 Free PMC article.
-
Quantitative perturbation-phenotype maps reveal nonlinear responses underlying robustness of PAR-dependent asymmetric cell division.PLoS Biol. 2024 Dec 9;22(12):e3002437. doi: 10.1371/journal.pbio.3002437. eCollection 2024 Dec. PLoS Biol. 2024. PMID: 39652540 Free PMC article.
-
Dynein modifiers in C. elegans: light chains suppress conditional heavy chain mutants.PLoS Genet. 2007 Aug;3(8):e128. doi: 10.1371/journal.pgen.0030128. PLoS Genet. 2007. PMID: 17676955 Free PMC article.
References
-
- Betschinger, J., and J. A. Knoblich, 2004. Dare to be different: asymmetric cell division in Drosophila, C. elegans and vertebrates. Curr. Biol. 14: R674–R685. - PubMed
-
- Boyd, L., S. Guo, D. Levitan, D. T. Stinchcomb and K. J. Kemphues, 1996. PAR-2 is asymmetrically distributed and promotes association of P granules and PAR-1 with the cortex in C. elegans embryos. Development 122: 3075–3084. - PubMed
-
- Cheeks, R. J., J. C. Canman, W. N. Gabriel, N. Meyer, S. Strome et al., 2004. C. elegans PAR proteins function by mobilizing and stabilizing asymmetrically localized protein complexes. Curr. Biol. 14: 851–862. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
